Vivchenni . के साथ शुभकामनाएँ न्यूक्लिक एसिडऔर प्रोटीन के जैवसंश्लेषण को निम्न विधियों के विकास के लिए लाया गया था, जो चिकित्सा, मजबूत स्थिति और अन्य दीर्घाओं में बहुत व्यावहारिक महत्व का हो सकता है।
इसके अलावा, आनुवंशिक कोड और मंदी की जानकारी को बचाने और लागू करने के बुनियादी सिद्धांतों के रूप में, बहरे कुट में जैश के आणविक जीव विज्ञान के विकास, कोई तरीके नहीं थे, क्योंकि उन्हें जीन में हेरफेर करने, उन्हें देखने और बदलने की अनुमति थी। . इन विधियों का उद्भव 1970-1980 के दशक में शुरू हुआ। त्से ने विज्ञान के चक्र के विकास के लिए एक थकाऊ पद दिया, याक और आज विकास की अवधि का अनुभव कर रहे हैं। Nasampered, व्यक्तिगत जीन के चयन और अन्य जीवों (आणविक क्लोनिंग और ट्रांसजेनेसिस, PLR) की कोशिकाओं में उनके परिचय के साथ-साथ जीन (डीएनए और आरएनए अनुक्रमण) में न्यूक्लियोटाइड के अनुक्रम को निर्धारित करने के तरीके हैं। नीचे, इन विधियों की रिपोर्ट द्वारा समीक्षा की जाएगी। आइए सबसे सरल बुनियादी विधि - वैद्युतकणसंचलन से शुरू करें और फिर तह विधियों पर आगे बढ़ें।
डीएनए इलेक्ट्रोफोरेसिस
यह डीएनए के साथ काम करने का मूल तरीका है, जिसे एक ही समय में व्यावहारिक रूप से अन्य सभी तरीकों से विकसित किया जाता है ताकि आवश्यक अणुओं को देखा जा सके और परिणामों का विश्लेषण किया जा सके। अधिक डीएनए अंशों के लिए, जेल वैद्युतकणसंचलन की विधि का उपयोग लंबे समय तक किया जाता है। डीएनए एक एसिड है, अणु अतिरिक्त फॉस्फोरिक एसिड को दूर करते हैं, याक एक प्रोटॉन को विभाजित करते हैं और एक नकारात्मक चार्ज प्राप्त करते हैं (चित्र 1)।
टॉम इन विद्युत क्षेत्रडीएनए अणु एनोड में गिर जाते हैं - एक सकारात्मक चार्ज इलेक्ट्रोड। Tse vіdbuvaєtsya rozchinі elektrolіtіv में, scho प्रतिशोध आयन-वाहक चार्ज, zavdjaki क्यों tsey rozchin को स्ट्रम करने के लिए। टुकड़ों को विभाजित करने के लिए, पॉलिमर (एग्रोस या पॉलीएक्रिलामाइड) से एक क्षारीय जेल बनाया जाता है। डीएनए अणु नवीनतम समय में बदबू से अधिक "खो जाते हैं", और इसलिए पाए गए अणु सबसे अधिक ढह जाते हैं, और सबसे छोटे - सबसे अधिक (चित्र 2)। पहले, वैद्युतकणसंचलन के बाद, जेल को बार्नाकल के साथ इलाज किया जाता है, जो पराबैंगनी प्रकाश में डीएनए और फ्लोरोसेंट से बंधता है, और जेल में धुंध की तस्वीर लेता है (div। चित्र 3)। डीएनए अंशों की लंबाई को निर्दिष्ट करने के लिए, उनकी तुलना एक मार्कर से की जानी चाहिए - एक ही जेल पर समानांतर में लागू मानक लंबाई में टुकड़ों का एक सेट (चित्र 4)।



डीएनए के साथ काम करने के लिए सबसे महत्वपूर्ण उपकरण एंजाइम हैं जो जीवित कोशिकाओं में डीएनए के परिवर्तन को प्रभावित करते हैं: डीएनए पोलीमरेज़, डीएनए बंधन और प्रतिबंध एंडोन्यूक्लिज़, या प्रतिबंध एंडोन्यूक्लिज़। डीएनए पोलीमरेज़डीएनए के zdіysnyuyut मैट्रिक्स संश्लेषण, जो नमूने में डीएनए को दोहराने की अनुमति देता है। डीएनए लिगेज zshivayut आपस में DNA के अणु या zalіkovuyut उन्हें तोड़ते हैं। प्रतिबंध एंडोन्यूक्लाइजेस, या प्रतिबंधित करना, गायन अनुक्रमों के बाद डीएनए अणुओं को अलग करें, जो मुख्य डीएनए द्रव्यमान से पूरे टुकड़े की कल्पना करने की अनुमति देता है। कुछ मामलों में क्यूई अंशों को जीन के रूप में पुन: उत्पन्न किया जा सकता है।
प्रतिबंधित करना
प्रतिबंध एंजाइमों द्वारा नियंत्रित, सममित और विकसित अनुक्रम ऐसे अनुक्रम के बीच में या डीएनए के दोनों स्ट्रैंड में एक ही स्थान पर पाए जा सकते हैं। विभिन्न प्रकार के प्रतिबंध की योजना को अंजीर में दिखाया गया है। 1. पहले प्रकार का नाम "बेवकूफ" kіnci है, जबकि दूसरे में "चिपचिपा" kіnci है। तल के "चिपचिपे" सिरों के समय में, भाले दूसरे की तुलना में छोटे दिखाई देते हैं, एक एकल-फंसे नाव को एक सममित अनुक्रम के साथ स्थापित किया जाता है, दोनों सिरों पर समान होता है, जो व्यवस्थित होते हैं।
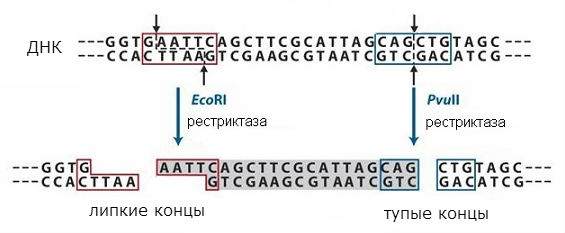
अंतिम क्रम वही होगा जब विभाजित किया जाता है कि डीएनए किसी दिए गए प्रतिबंध एंजाइम द्वारा दिया गया है या नहीं और फिर से जुड़ सकता है, टुकड़े पूरक अनुक्रम हो सकते हैं। अतिरिक्त डीएनए-बंधाव के लिए Їx को सीवन किया जा सकता है और एक एकल अणु ले सकते हैं। इस तरह, दो अलग-अलग डीएनए के टुकड़े एकजुट हो सकते हैं और तथाकथित ले सकते हैं पुनः संयोजक डीएनए. यह दृष्टिकोण आणविक क्लोनिंग की विधि पर आधारित है, जो व्यक्तिगत जीन के चयन को कोशिकाओं में पेश करने की अनुमति देता है, ताकि वे जीन प्रोटीन में कोडिंग स्थापित कर सकें।
आणविक क्लोनिंग
आणविक क्लोनिंग में दो डीएनए अणु होते हैं - एक इंसर्ट, क्या जीन का बदला लेना है, क्या दबाना है, और वेक्टर- डीएनए कि भूमिका है। अतिरिक्त एंजाइमों के लिए वेक्टर में सम्मिलन "सिलना" होता है, एक नया, पुनः संयोजक डीएनए अणु लेता है, फिर यह अणु मास्टर कोशिकाओं को पारित किया जाता है, और कोशिकाएं जीवन देने वाले माध्यम पर उपनिवेश स्थापित करती हैं। एक कॉलोनी एक क्लिटिन की संतान होती है, यानी एक क्लोन, एक कॉलोनी के सभी क्लिटिन आनुवंशिक रूप से समान होते हैं और एक ही पुनः संयोजक डीएनए को प्रतिस्थापित करते हैं। इसलिए, "आणविक क्लोनिंग" शब्द का अर्थ है कि डीएनए के एक टुकड़े का बदला लेने के लिए, हमें बदनाम करने के लिए, क्लिटिन का क्लोन छीन लिया जाता है। इसके अलावा, एक उपनिवेश के रूप में, सम्मिलन का बदला लेने के लिए, हमें बदनाम करने के लिए, ओट्रीमनी, विभिन्न तरीकों से सम्मिलन को चिह्नित करना संभव है, उदाहरण के लिए, सटीक अनुक्रम निर्धारित करने के लिए। इसके अलावा, कोशिकाएं प्रोटीन को कंपन कर सकती हैं, जो एक इंसर्ट द्वारा एन्कोडेड होते हैं, ताकि वे एक कार्यात्मक जीन का बदला ले सकें।
क्लिटिन के पुनः संयोजक अणु की शुरूआत के साथ, इन क्लिटिन का आनुवंशिक परिवर्तन देखा जाता है। परिवर्तन- जीव के जीव को निवास के माध्यम से मुक्त डीएनए अणु प्राप्त करने और उनके जीनोम में नवोदित होने की प्रक्रिया, जो इसके लिए नए क्षयकारी संकेतों के ऐसे क्लिटिन की उपस्थिति का कारण बनती है, जो जीव से जुड़ी होती है- डीएनए का दाता। उदाहरण के लिए, यदि प्रतिरोध जीन को एंटीबायोटिक एम्पीसिलीन में बदलने के लिए एक अणु पेश किया गया था, तो इस उपस्थिति में रूपांतरित बैक्टीरिया विकसित होंगे। परिवर्तन से पहले, एम्पीसिलीन ने अपनी मृत्यु का कारण बना, ताकि रूपांतरित कोशिकाओं में, एक नया संकेत पैदा हो।
वेक्टर
अधिकारियों की एक श्रृंखला की मां के लिए वेक्टर दोषी है:
सबसे पहले, डीएनए अणु स्पष्ट रूप से छोटा है, जिसे हेरफेर करना आसान है।
दूसरे तरीके से, अनुक्रम में डीएनए को बचाने और गुणा करने के लिए, यह उसी अनुक्रम का बदला लेने की जिम्मेदारी है, जो प्रतिकृति (प्रतिकृति का कोब, या प्रतिकृति की उत्पत्ति) सुनिश्चित करता है।
तीसरा, आप इसे नीचे ले जा सकते हैं मार्कर जीनक्या सुरक्षित तरीका है, केवल एक शांत भगशेफ, एक याक में एक वेक्टर का सेवन करने के बाद। एंटीबायोटिक दवाओं के प्रतिरोध के जीन के लिए कॉल करें - यहां तक कि एंटीबायोटिक दवाओं की उपस्थिति में भी सभी कोशिकाएं, ताकि वेक्टर का बदला न लेने के लिए, मर जाएं।
जीवाणु कोशिकाओं में जीनों की क्लोनिंग सबसे अधिक बार की जाती है, क्योंकि दुर्गंध को विकसित करना और तेजी से गुणा करना आसान होता है। बैक्टीरिया में, एक बड़ा आणविक डीएनए अणु होता है, आवश्यक बैक्टीरिया जीन - जीवाणु गुणसूत्र से बदला लेने के लिए न्यूक्लियोटाइड जोड़े के लाखों जोड़े होते हैं। कुछ बैक्टीरिया में क्रीमियन बैक्टीरिया डीएनए रिंगों के छोटे (कुछ हजार बेस पेयर) पाए जाते हैं, जिन्हें कहा जाता है प्लास्मिड(रेखा चित्र नम्बर 2)। मुख्य डीएनए की तरह, न्यूक्लियोटाइड के अनुक्रम को बदलने के लिए बदबू आती है, जो डीएनए प्रतिकृति (ओरी) सुनिश्चित करता है। प्लास्मिड मुख्य (गुणसूत्र) डीएनए में स्वतंत्र रूप से दोहराते हैं, यही वजह है कि क्लिटिनिड्स की बड़ी संख्या में प्रतियां होती हैं। इनमें से कई प्लास्मिड एंटीबायोटिक प्रतिरोध के जीन ले जाते हैं, जो क्लिटिन को संक्रमित करने की अनुमति देता है, जो प्लास्मिड को उत्कृष्ट क्लिटिन के रूप में ले जाता है। प्लास्मिड जो दो जीन ले जाते हैं, जो दो एंटीबायोटिक दवाओं के लिए प्रतिरोध प्रदान करते हैं, उदाहरण के लिए, टेट्रासाइक्लिन और एपमिसिलिन, सबसे अधिक बार पाए जाते हैं। ऐसे प्लास्मिड डीएनए, जीवाणु के मुख्य गुणसूत्र के विभिन्न प्रकार के डीएनए की कल्पना करने के लिए एक सरल विधि स्थापित करें।

ट्रांसजेनेसिस का महत्व
एक जीव से दूसरे जीव में जीन का स्थानांतरण ट्रांसजेनेसिस, साथ ही संशोधित जीव - ट्रांसजेनिक्स. सूक्ष्मजीव की कोशिकाओं में जीन को स्थानांतरित करने की विधि से, चिकित्सा आवश्यकताओं के लिए पुनः संयोजक प्रोटीन की तैयारी, ज़ोक्रेमा, मानव प्रोटीन समाप्त हो जाते हैं, जो प्रतिरक्षा चयापचय का कारण नहीं बनते हैं - इंटरफेरॉन, इंसुलिन और अन्य प्रोटीन हार्मोन, टीके। अधिक मुड़ी हुई अवस्थाओं में, यदि प्रोटीन का संशोधन केवल यूकेरियोट्स के क्लिटिन में सही ढंग से किया जाता है, तो ट्रांसजेनिक क्लिटिन संस्कृतियों, या ट्रांसजेनिक जीवों, ज़ोक्रेमा, पतलेपन (हम किज़ के लिए सबसे पहले हैं) को रोपना संभव है, जैसा कि हम देखते हैं दूध में आवश्यक प्रोटीन, अन्यथा रक्त से प्रोटीन दिखाई देता है। तो एंटीबॉडी, रक्त के कारक और अन्य प्रोटीन को हटा दें। ट्रांसजेनेसिस विधि द्वारा लिया गया सांस्कृतिक रोज़लिनी, जड़ी-बूटियों के लिए stіykі और shkіdnіkіv yakі mayut innshі korisnі प्राधिकरण। ट्रांसजेनिक सूक्ष्म जीवों की मदद के लिए, वे सीवर को शुद्ध करते हैं और उलझाव से लड़ते हैं, ट्रांसजेनिक रोगाणुओं को विकसित करते हैं, जो नेफ्था को विघटित कर सकते हैं। क्रीमिया, ट्रांसजेनिक प्रौद्योगिकियां अपरिहार्य हैं वैज्ञानिक उपलब्धियां- जीव विज्ञान का विकास आज जीन को संशोधित और स्थानांतरित करने के लिए नियमित zastosuvannya विधियों के बिना अकल्पनीय है।
आणविक क्लोनिंग तकनीक
आवेषण
किसी भी जीव से एक जीन को हटाने के लिए, सभी गुणसूत्र डीएनए को देखा जाता है और एक या दो प्रतिबंध एंजाइमों के साथ विभाजित किया जाता है। एंजाइमों को इस तरह से चुना जाता है कि बदबू हमारे लिए सिकेवियार जीन को नहीं काटती है, बल्कि इसके किनारों के साथ कट को तोड़ती है, और प्लास्मिड डीएनए में वे प्रतिरोध जीन में से एक में 1 कट तोड़ते हैं, उदाहरण के लिए, एम्पीसिलीन को।
आणविक क्लोनिंग प्रक्रिया में निम्नलिखित चरण शामिल हैं:
विभाजन और सिलाई - एक एकल पुनः संयोजक अणु के सम्मिलन और वेक्टर से निर्माण।
परिवर्तन - कोशिकाओं में एक पुनः संयोजक अणु की शुरूआत।
चयन - चयनित क्लिटिन, याक ने वेक्टर को डालने से हटा दिया।
काटने और सिलाई
प्लास्मिड डीएनए को उसी प्रतिबंध एंजाइम द्वारा संसाधित किया जाता है, और इसे एक रैखिक अणु में बदल दिया जाता है, क्योंकि इस तरह के प्रतिबंध एंजाइम का चयन किया जाता है, ताकि 1 उद्घाटन प्लास्मिड में पेश किया जा सके। डीएनए अंशों के सभी सिरों के परिणामस्वरूप वही चिपचिपे सिरे सामने आते हैं। कम तापमान पर, किंक एक अवसाद में बदल जाते हैं और वे डीएनए लिगेज (डिव। चित्र 3) के साथ क्रॉस-लिंक्ड होते हैं।

विभिन्न गोदामों के क्रोमोसोमल डीएनए की रकम निकालें: उनमें से कुछ बैक्टीरिया डीएनए से प्राप्त क्रोमोसोमल डीएनए के पूरे डीएनए अनुक्रम को बदल देंगे, अन्य एक बार में क्रोमोसोमल डीएनए के टुकड़ों को जोड़ देंगे, और तीसरा क्रोमोसोमल डीएनए या डिमर के पूरे अनुक्रम को बदल देगा। (चित्र 4)।

परिवर्तन
मुझे अपनी राशि खर्च करने दो आनुवंशिक परिवर्तनबैक्टीरिया, ताकि प्लास्मिड के साथ प्रतिशोध न करें। परिवर्तन- जीव के जीव को निवास के माध्यम से मुक्त डीएनए अणु प्राप्त करने और उनके जीनोम में नवोदित होने की प्रक्रिया, जो इसके लिए नए क्षयकारी संकेतों के ऐसे क्लिटिन की उपस्थिति का कारण बनती है, जो जीव से जुड़ी होती है- डीएनए का दाता। त्वचा कोशिका में, यह प्रवेश कर सकता है और गुणा कर सकता है, केवल एक प्लाज्मिड होता है। ऐसी कोशिकाओं को एक कठोर जीवन देने वाले माध्यम पर रखा जाता है, जिसमें एंटीबायोटिक टेट्रासाइक्लिन स्थित होता है। क्लिटिनी, याक ने प्लास्मिड का उपभोग नहीं किया, जो माध्यम नहीं बढ़ता है, और क्लिटिनी, जो प्लास्मिड ले जाती है, कॉलोनियां स्थापित करती है, उनमें से त्वचा जेड में पैच क्लिटिनी, टोबो में से एक से कम हैं। कॉलोनी की सभी कोशिकाओं में एक ही प्लास्मिड होता है (div। चित्र 5)।

चयन
डाली लागत ज़ावदन्न्या केवल क्लिटिन देखते हैं, एक तरह से डालने से वेक्टर का सेवन करते हैं, और अपने प्रकार के क्लिटिन को संशोधित करते हैं, जो केवल वेक्टर को बिना सम्मिलित किए ले जाते हैं, या ज़ोवसिम वेक्टर नहीं ले जाते हैं। आवश्यक ग्राहकों के चयन की पूरी प्रक्रिया कहलाती है चयन. किसके लिए रुकें चयनात्मक मार्कर- वेक्टर के गोदाम में एंटीबायोटिक दवाओं के आनुवंशिक प्रतिरोध को बुलाओ, कि चयनात्मक मीडियाएंटीबायोटिक्स या अन्य भाषण का बदला कैसे लें, चयन कैसे सुनिश्चित करें
हमारे मामले में, एम्पीसिलीन की उपस्थिति में बढ़ने वाली कॉलोनियों की कोशिकाओं को दो माध्यमों में विभाजित किया जाता है: पहले एक में - एम्पीसिलीन, और दूसरे में - टेट्रासाइक्लिन। कॉलोनियां, जो केवल प्लास्मिड पर हमला करती हैं, मीडिया और कॉलोनियों दोनों पर विकसित होती हैं, जिनमें प्लास्मिड में टेट्रासाइक्लिन के साथ माध्यम पर क्रोमोसोमल डीएनए होता है, विकसित नहीं होते हैं (चित्र 5)। उनमें से, हमारे लिए जीन का चयन करने, पर्याप्त संख्या में बढ़ने और प्लास्मिड डीएनए को देखने के लिए विशेष विधियों का उपयोग किया जाता है। इसके साथ, उन्हीं प्रतिबंधों की मदद से, जो पुनः संयोजक डीएनए की वापसी के साथ विजयी हुए थे, वे एक व्यक्तिगत जीन की कल्पना करने में सक्षम थे जो क्लीट करता है। नई शक्तियों को हटाने या आवश्यक प्रोटीन के संश्लेषण के लिए किसी भी जीव में पेश किए गए न्यूक्लियोटाइड के अनुक्रम के पदनाम के लिए इस जीन का डीएनए विजयी हो सकता है। जीन देखने की इस विधि को कहते हैं आणविक क्लोनिंग.
फ्लोरोसेंट बिल्स
पिछले यूकेरियोटिक जीवों के मामले में आनुवंशिक मार्कर के रूप में, फ्लोरोसेंट प्रोटीन को मैन्युअल रूप से बदलना भी संभव है। पहला फ्लोरोसेंट प्रोटीन जीन, हरित प्रोटीन जो प्रतिदीप्त होता है (हरा प्रतिदीप्त प्रोटीन, GFP)हमने विभिन्न मॉडल जीवों में जेलिफ़िश एक्यूओरिया विक्टोरिया और इनोक्यूलेशन को देखा है (डिव। चित्र 6)। 2008 में, O. Simomura, M. Chalfi, और R. Tsien को प्रोटीन की आपूर्ति पर उनके काम के लिए नोबेल पुरस्कार से सम्मानित किया गया था।

फिर हमने अन्य फ्लोरोसेंट प्रोटीन के जीन देखे - लाल, नीला, पीला। भूरे रंग की शक्तियों वाले प्रोटीन को दूर करने के लिए क्यूई जीन को टुकड़े-टुकड़े संशोधित किया गया था। फ्लोरोसेंट प्रोटीन की बहुमुखी प्रतिभा को अंजीर में दिखाया गया है। चित्रा 7 विभिन्न फ्लोरोसेंट प्रोटीन के जीन का बदला लेने के लिए बैक्टीरिया के साथ एक पेट्री डिश दिखाता है।

फ्लोरोसेंट प्रोटीन का आसव
एक फ्लोरोसेंट प्रोटीन के जीन को किसी अन्य प्रोटीन के जीन के साथ क्रॉसलिंक किया जा सकता है, फिर अनुवाद के दौरान एक एकल प्रोटीन का उत्पादन होता है - एक ट्रांसलेशनल प्रोटीन, या विलय(फ्यूजन प्रोटीन), जो फ्लोरोसेंट है। इस तरह के एक संस्कार में, आप, उदाहरण के लिए, स्थानीय (रोज़ताशुवन्न्या) कर सकते हैं कि क्या गोरे हैं, क्या कहें, क्लिटिन में, उनका आंदोलन। एकल प्रकार के क्लिटिन में फ्लोरोसेंट प्रोटीन की अतिरिक्त अभिव्यक्ति के लिए, इन प्रकार की कोशिकाओं को एक समृद्ध क्लिटिनस जीव में चिह्नित करना संभव है (div। चित्र 8 - एक भालू का मस्तिष्क, जिसमें वातावरण में न्यूरॉन्स के अलग-अलग रंग हो सकते हैं) सिंगफुल फ्लोरोसेंट फ्लोरेसेंस के रेचनिड्स के लिए)। फ्लोरोसेंट प्रोटीन आधुनिक आणविक जीव विज्ञान में एक अनिवार्य उपकरण हैं।

पीएलआर
जीन को हटाने की एक अन्य विधि कहलाती है पोलीमरेज़ लैनज़ग रिएक्शन (PLR). यह डीएनए पोलीमरेज़ की पूरक स्ट्रैंड के बाद एक और डीएनए स्ट्रैंड प्राप्त करने की क्षमता पर आधारित है, जैसा कि डीएनए प्रतिकृति के दौरान कोशिकाओं में होता है।
इस विधि में प्रतिकृति के सिल पर बिंदु डीएनए के दो छोटे टुकड़ों द्वारा निर्धारित किए जाते हैं, जिन्हें कहा जाता है बीज,या प्राइमरों. क्यूई प्राइमर दो डीएनए लेंस पर जीन के सिरों, scho tsіkavit के पूरक हैं। गुणसूत्र डीएनए का पिछला भाग, जिसे देखने के लिए जीन की आवश्यकता होती है, प्राइमरों के साथ मिलाया जाता है और लगभग 99 C तक गर्म किया जाता है। पानी का उत्पादन करने के लिए जब तक कि पानी के लिंक टूट नहीं जाते और डीएनए स्ट्रैंड अलग नहीं हो जाते। इसके बाद, तापमान 50-70 डिग्री सेल्सियस (बीजारोपण के क्रम के कारण जमा) तक कम हो जाता है। इन दिमागों में, प्राइमर क्रोमोसोमल डीएनए की पूरक कोशिकाओं से जुड़े होते हैं, जिससे सही सबफोल्डिंग हेलिक्स (div। चित्र 9) स्थापित होता है। उसके बाद, डीएनए संश्लेषण के लिए आवश्यक सभी कोटिरियो न्यूक्लियोटाइड्स का योग जोड़ें, वह डीएनए पोलीमरेज़। एंजाइम podzhuє भड़काना, भड़काना, टोबो के लगाव की जगह के डबल-असहाय डीएनए होने के नाते। जीन के अंत में, सिंगल लैंसेट क्रोमोसोम अणु के अंत तक। 
अब, अगर मैं योग को फिर से गर्म करता हूं, तो क्रोमोसोम और लांसर्स के नए संश्लेषण में वृद्धि होगी। ठंडा होने के बाद, अंकुर उनके पास वापस आ जाते हैं, क्योंकि उन्हें बहुत अधिक मात्रा में लिया जाता है (div। चित्र 10)।

नए संश्लेषित लेंसों पर, बदबू अंत तक नहीं आती है, जहां से पहला संश्लेषण शुरू हुआ था, लेकिन प्रोटोलिथिक के लिए, क्योंकि डीएनए लेंस एंटीपैरेलल हैं। इसलिए, ऐसे लेंसों पर संश्लेषण के एक अन्य चक्र में, केवल अनुक्रम प्राप्त होगा, जो कि जीन के समान है (div। चित्र 11)।

इस विधि में, थर्मोफिलिक बैक्टीरिया से डीएनए पोलीमरेज़ को टीका लगाया जाता है, इसे उबाला जा सकता है और लगभग 70-80 C के तापमान पर संसाधित किया जा सकता है, और इसमें स्कोरेज़ जोड़ने की आवश्यकता नहीं होती है, लेकिन यह कोब में एक संदेश जोड़ने के लिए पर्याप्त है। उसी क्रम में हीटिंग और कूलिंग की प्रक्रियाओं को दोहराकर, हम त्वचा चक्र में अनुक्रमों की संख्या पर जीत हासिल कर सकते हैं, obmezheniya z dvoh kintsіv प्राइमिंग (div। चित्र 12)।

ऐसे लगभग 25 चक्रों के बाद जीन की प्रतियों की संख्या कमोबेश दस लाख गुना बढ़ जाएगी। इतनी मात्रा को टेस्ट ट्यूब में पेश किए गए क्रोमोसोमल डीएनए से आसानी से पुनर्प्राप्त किया जा सकता है और विभिन्न उद्देश्यों के लिए विजयी हो सकता है।
डीएनए श्रृंखला बनाना
एक अन्य महत्वपूर्ण उपलब्धि डीएनए में न्यूक्लियोटाइड के अनुक्रम को निर्धारित करने के तरीकों का विकास है। डीएनए श्रृंखला बनाना(अंग्रेजी अनुक्रम से - अनुक्रम)। इसके लिए वर्णित विधियों में से किसी एक द्वारा जीन के शुद्धतम डीएनए का चयन करना आवश्यक है। फिर डीएनए लेंसों को गर्म करके अलग किया जाता है और उन्हें रेडियोधर्मी फास्फोरस या फ्लोरोसेंट लेबल के साथ लेबल किया जाता है। सम्मान प्राप्त करने के लिए, जो एक उत्पीड़न द्वारा लिया जाता है, एक लांसर का पूरक। फिर हम डीएनए पोलीमरेज़ और 4 न्यूक्लियोटाइड का योग जोड़ते हैं। इस तरह की राशि को 4 भागों में विभाजित किया जाता है और न्यूक्लियोटाइड्स में से एक को त्वचा में जोड़ा जाता है, संशोधन करता है ताकि वाइन के डीऑक्सीराइबोज का तीसरा परमाणु हाइड्रॉक्सिल समूह का बदला न ले सके। यदि ऐसे न्यूक्लियोटाइड को डीएनए लेंस के संश्लेषण में शामिल किया जाता है, तो इसे जारी रखा जा सकता है, क्योंकि पोलीमरेज़ कहीं भी न्यूक्लियोटाइड को आगे बढ़ाते हैं। इसलिए, ऐसे न्यूक्लियोटाइड को शामिल करने के बाद डीएनए संश्लेषण काट दिया जाता है। इस तरह के न्यूक्लियोटाइड्स, डाइडॉक्सिन्यूक्लियोटाइड्स के शीर्षक, काफी कम, कम दिए जाते हैं, इसलिए लैंसेट यूरविच कम आम है और अन्य क्षेत्रों में त्वचा लैंसेट में है। नतीजतन, विभिन्न लंबाई के लेंसों का योग निकलता है, त्वचा के सिरों पर एक ही न्यूक्लियोटाइड होता है। इस क्रम में, लैंसलेट की लंबाई अनुक्रम में न्यूक्लियोटाइड की संख्या से मेल खाती है, जिसे दोहराया जाता है, उदाहरण के लिए, जैसा कि हमारे पास एडेनिल डिडॉक्सिन्यूक्लियोटाइड है, और लैंसलेट की ओट्रीमनी 2, 7 की लंबाई से छोटी थी और 12 न्यूक्लियोटाइड्स, जिसका अर्थ है जीन में दूसरे, सातवें और बारहवें स्थान पर। अतिरिक्त वैद्युतकणसंचलन के लिए मेंहदी द्वारा लेंस के ओट्रिमैन योग को आसानी से अलग किया जा सकता है, और एक्स-रे तैराकों पर रेडियोधर्मिता द्वारा लेंस के संश्लेषण का पता लगाया जा सकता है (div। चित्र 10)।

एक तस्वीर सामने आती है, नीचे की तरफ एक छोटा सा इशारा किया जाता है, जिसे रेडियो ऑटोग्राफ कहा जाता है। नए तल के साथ ऊपर की ओर दौड़ते हुए और त्वचा क्षेत्र के स्तंभों के ऊपर के पत्र को पढ़ते हुए, हम न्यूक्लियोटाइड्स के अनुक्रम को हटा देते हैं, ऑटोग्राफ पर दाहिने हाथ की ओर इशारा करते हुए। यह पता चला कि संश्लेषण न केवल डिडॉक्सिन्यूक्लियोटाइड्स पर आधारित है, बल्कि न्यूक्लियोटाइड्स पर भी आधारित है, जिसमें, तीसरे स्थान पर, सर्किट में एक रासायनिक समूह होता है, उदाहरण के लिए, एक फ्लोरोसेंट खलिहान। यदि आप अपने खलिहान के साथ त्वचा के न्यूक्लियोटाइड को चिह्नित करते हैं, तो ज़ोन, जो लेंस के संश्लेषण के पृथक्करण के दौरान हटा दिए जाते हैं, एक गुलाबी रोशनी से चमकते हैं। यह आपको सभी न्यूक्लियोटाइड के लिए एक समय में एक नमूने में प्रतिक्रिया करने और कबूतर द्वारा लैंसेट के स्ट्रिप्स को अलग करने की अनुमति देता है, न्यूक्लियोटाइड के लिए रंगों की पहचान करता है (div। चित्र 11)।

इस तरह के तरीकों ने एक दर्जन जीनों की तरह अनुक्रमों को नामित करना और पूरे जीनोम को पढ़ना संभव बना दिया। वर्तमान समय में, जीनों में न्यूक्लियोटाइड के अनुक्रम निर्दिष्ट करने के लिए और अधिक आधुनिक तरीके विकसित किए गए हैं। जैसे मानव भाप जीनोम को एक महान अंतरराष्ट्रीय संघ द्वारा 12 साल में विधि द्वारा प्रेरित किया गया था, दूसरे के साथ, तीन साल में, फिर इसे एक महीने में तोड़ा जा सकता था। त्से आपको किसी व्यक्ति के विद्वान को समृद्ध बीमारी में स्थानांतरित करने की अनुमति देता है, और फिर खो जाने के लिए पर्याप्त समय तक जीने की अनुमति देता है।
XX सदी के 40 के दशक के सिल पर लगभग जैव रसायन, बायोफिज़िक्स, जेनेटिक्स, साइटोकेमिस्ट्री, माइक्रोबायोलॉजी और वायरोलॉजी में समृद्ध विकास का विकास। vpritul ने आणविक स्तर पर जीवन की घटनाओं के उद्भव का नेतृत्व किया। इन विज्ञानों द्वारा एक ही समय में और विभिन्न पक्षों से प्राप्त सफलताओं ने इस तथ्य को ध्यान में लाया कि शरीर की मुख्य आवश्यक प्रणालियाँ आणविक स्तर पर कार्य करती हैं, और इन विज्ञानों में आगे की प्रगति के विकास के कारण है। अणुओं के जैविक कार्य जो शरीर के शरीर को स्थापित करते हैं। क्लिटीना से विघटन, पारस्परिक परिवर्तन और प्रजनन, साथ ही साथ ऊर्जा और सूचना का आदान-प्रदान, जो इसके साथ आता है। तो, इन जैविक विषयों, रसायन विज्ञान और विनाइल भौतिकी के आधार पर, आणविक जीव विज्ञान एक नई चुनौती है।
Vіdmin Vigo Bіokhimії, Uvagi, आणविक आणविक Bischeni क्षेत्र पर, Vivchennі Structuri पर पचता है, funki-bіlkіv bіopolіvіv - bіlkіv, ऐसे न्यूक्लिक एसिड, खुद Prosikanniy Richai के राजकुमार, और Bіosynthesis के अन्य - bіosynthesis के कार्य। यह स्पष्ट था कि आणविक जीव विज्ञान और जैव रसायन, आनुवंशिकी के संबंधित प्रभागों, सूक्ष्म जीव विज्ञान और वायरोलॉजी के बीच अंतर करना असंभव है।
आणविक जीव विज्ञान का आरोप अनुसंधान के नए तरीकों के विकास से निकटता से संबंधित था, जो पहले से ही अन्य शाखाओं में देखा गया था। इलेक्ट्रॉन माइक्रोस्कोपी में कई विकास और सूक्ष्म प्रौद्योगिकी में अन्य तरीकों ने सेलुलर तत्वों के विभाजन के 50 के तरीकों के विकास में एक महान भूमिका निभाई। बदबू को विभेदक सेंट्रीफ्यूजेशन (ए। क्लाउड, 1954) के संपूर्ण तरीकों पर आधारित किया गया था। इस घंटे तक, वे पहले से ही बायोपॉलिमर के उस अंश को देखने के सर्वोत्तम तरीकों को खोजने की कोशिश कर रहे थे। हियर टू लेट, ज़ोक्रेमा, ए. टिसेलियस द्वारा प्रस्तावना (1937; नोबेल पुरस्कार, 1948) अतिरिक्त वैद्युतकणसंचलन के लिए प्रोटीन के विभाजन की विधि, न्यूक्लिक एसिड के अवलोकन और शुद्धिकरण की विधि (ई। के, ए। डाउन्स, एम। सेवाग, ए मिर्स्की और आईएनजी।) उसी समय, विभिन्न प्रयोगशालाओं (ए। मार्टिन और आर। सिंग, 1941; नोबेल पुरस्कार, 1952) में विभिन्न तरीकों और क्रोमैटोग्राफिक विश्लेषण विकसित किए गए थे, और यह पूरी तरह से किया गया था।
एक्स-रे विवर्तन विश्लेषण का उपयोग करके बायोपॉलिमर की संरचना को समझने में एक अमूल्य सेवा। एक्स-रे विवर्तन विश्लेषण के मूल सिद्धांतों को किंग्स कॉलेज लंदन विश्वविद्यालय में डब्ल्यू। ब्रैग के शोध के तहत योगदानकर्ताओं के एक समूह द्वारा साझा किया गया था, जिसमें जे। बर्नल, ए। लोंड्सडेल, डब्ल्यू। एस्टबरी, जे। रॉबर्टसन और अन्य शामिल थे।
मॉस्को स्टेट यूनिवर्सिटी के प्रोफेसर ए.आर. Kizel, एक झटका मारा, जड़ लेने के बाद, यह पता चला कि प्रोटोप्लाज्म के आधार पर, यह एक विशेष प्रोटीन शरीर - प्लेट हो, जो निबिटो सभी सबसे महत्वपूर्ण संरचनात्मक और कार्यात्मक विशेषताओं को दर्शाता है। यह दिखाने के बाद कि प्लेटें एकमात्र प्रोटीन हैं, जो केवल myxomycetes में बढ़ती हैं, और फिर विकास के गायन चरण में, और वही स्थिर घटक - एक एकल कंकाल प्रोटीन - प्रोटोप्लाज्म में मौजूद नहीं होता है। अपने आप से, प्रोटोप्लाज्म की समस्या के विकास और प्रोटीन की कार्यात्मक भूमिका ने सही रास्ता अपनाया और इसके विकास के लिए जगह हासिल की। सेल के गोदाम भागों के रसायन विज्ञान के विकास को उत्तेजित करते हुए, किज़ेल की उपलब्धियों ने मान्यता का प्रकाश जीता।
शब्द "आणविक जीव विज्ञान" पहली बार अंग्रेजी क्रिस्टलोग्राफर, लीड्स विश्वविद्यालय के प्रोफेसर डब्ल्यू। एस्टबरी, विनिक, यमोविर्नो द्वारा 40 के दशक के कोब पर (1945 तक) गढ़ा गया था। 1930 के दशक में एस्टबरी द्वारा किए गए प्रोटीन और डीएनए के मुख्य एक्स-रे विवर्तन अध्ययन ने इन बायोपॉलिमर की माध्यमिक संरचना के एक और सफल गूढ़ रहस्य के आधार के रूप में कार्य किया। 1963 पी. जे. बर्नाल ने लिखा: "आपके लिए स्मारक सभी आणविक जीव विज्ञान द्वारा बनाया जाएगा - एक विज्ञान, वाइन का नामकरण और ठीक से सो रहा है" * अंग्रेजी पत्रिका "नेचर" ** में प्रकाशित डब्ल्यू। एस्टबरी के लेख "एक्स-रे विवर्तन की प्रगति" कार्बनिक और रेशेदार बीजाणुओं का विश्लेषण। एस्टबरी के हार्वे व्याख्यान (1950) ने संकेत दिया: मुझे थोड़ा ymovіrno चाहिए, कि मैं पहले प्रोपोनुवव योगो . 1950 की शुरुआत में ही एस्टबरी यह स्पष्ट हो गया था कि आणविक जीव विज्ञान मैक्रोमोलेक्यूल्स की संरचना और संरचना से ठीक आगे हो सकता है, जिसका विकास जीवित जीवों के कामकाज की समझ के लिए सर्वोपरि है।
* (बायोग्र मेम. साथियों रॉय। समाज, 1963, वी. 9, 29.)
** (डब्ल्यू टी एस्टबरी। कार्बनिक और फाइबर संरचनाओं के एक्स-रे विश्लेषण की प्रगति।- प्रकृति,। 1946, वी. 157, 121.)
*** (डब्ल्यू टी एस्टबरी। आण्विक जीवविज्ञान में एडवेंचर्स। थॉमस स्प्रिंगफील्ड, 1952, पी. 3.)
आण्विक जीव विज्ञान के सामने खड़े थे, प्रकाश में, वही नेता, जो और पूरे जीव विज्ञान के सामने, - मुख्य घटना के उस योग के दैनिक जीवन का ज्ञान, ऐसे ज़ोक्रेमा, जैसे स्पैडकोविस्ट और मामूली। आधुनिक आणविक जीव विज्ञान को जीवों की आनुवंशिक जानकारी के कार्यान्वयन के लिए ओटोजेनी के विभिन्न चरणों और पढ़ने के विभिन्न चरणों में जीन, पथ और तंत्र की संरचना और कार्य को समझने के लिए कहा गया है। वॉन ने उत्परिवर्तजन की प्रकृति और विकासवादी प्रक्रिया के आणविक आधार की व्याख्या करने के लिए जीन गतिविधि और सेलुलर भेदभाव के नियमन के सूक्ष्म तंत्र की खोज का आह्वान किया।
न्यूक्लिक एसिड की आनुवंशिक भूमिका की स्थापना
आणविक जीव विज्ञान के विकास के लिए, सबसे महत्वपूर्ण ऐसे निष्कर्षों की कम संख्या है। 1944 में अमेरिकी शोधकर्ता ओ. एवरी, के. मैकलियोड (नोबेल पुरस्कार, 1923) और एम. मैकार्थी ने दिखाया कि न्यूमोकोकी में देखे जाने वाले डीएनए अणुओं में परिवर्तनकारी गतिविधि हो सकती है। डीऑक्सीराइबोन्यूक्लिएज द्वारा डीएनए के हाइड्रोलिसिस के बाद, रूपांतरण गतिविधि अधिक स्पष्ट थी। टिम स्वयं इस पर पुनर्विचार करने वाले पहले व्यक्ति थे कि क्लिटिन में आनुवंशिक कार्य स्वयं डीएनए से संपन्न थे, न कि प्रोटीन के साथ।
न्याय के लिए, यह ध्यान दिया जाना चाहिए कि जीवाणु परिवर्तन की घटना एवर, मैकलियोड और मैककार्थ की गवाही से काफी पहले खोजी गई थी। 1928 में एफ। ग्रिफिट ने एक लेख प्रकाशित किया, जिसमें उन्होंने समझाया कि कैप्सुलेटेड वायरल स्ट्रेन के मारे गए कोशिकाओं के गैर-एनकैप्सुलेटिंग (गैर-एनकैप्सुलेटिंग) न्यूमोकोकी को जोड़ने के बाद, कोशिकाओं का योग चूहों के लिए घातक हो गया। इसके अलावा, न्यूमोकोकी की जीवित कोशिकाएं, जो पागल प्राणियों से संक्रमित से देखी जाती हैं, पहले से ही विषाणुजनित थीं और उनमें एक छोटा पॉलीसेकेराइड कैप्सूल था। टिम ने स्वयं अपने विवेक से दिखाया है कि मृत न्यूमोकोकल कोशिकाओं के कुछ घटकों के प्रभाव में, जीवाणुओं का गैर-एनकैप्सुलेटेड रूप एक कैप्सुलर-गठन विषाणु रूप में बदल जाता है। 16 वर्षों के बाद, एवरी, मैकलियोड और मैककार्ट ने न्यूमोकोकी को डीऑक्सीराइबोन्यूक्लिक एसिड से बदल दिया और दिखाया कि डीएनए में ही परिवर्तनकारी गतिविधि है (डिव। स्प्लिट 7 और 25)। इस मत के महत्व का पुनर्मूल्यांकन करना महत्वपूर्ण है। इसने दुनिया की समृद्ध प्रयोगशालाओं में न्यूक्लिक एसिड के उत्पादन को प्रेरित किया और खुद वैज्ञानिकों का ध्यान डीएनए पर केंद्रित करने के लिए मजबूर किया।
50 के दशक के कोब पर एवरी, मैकलियोड और मैकार्थी द्वारा निर्देशित, इसने पहले से ही इस तथ्य के लिए बहुत सी प्रत्यक्ष और अप्रत्यक्ष श्रद्धांजलि जमा की है कि न्यूक्लिक एसिड जीवन में एक महत्वपूर्ण भूमिका निभाते हैं और एक आनुवंशिक कार्य करते हैं। त्से पर, ज़ोक्रेमा, क्लिटिन में डीएनए स्थानीयकरण की प्रकृति और उन लोगों के बारे में आर। वेंड्रेल (1948) दिखा रहा है, जहां डीएनए को क्लिटिन पर सख्ती से पोस्टियनो पर रखा गया है और प्रशंसनीयता की डिग्री के साथ सहसंबद्ध है: अगुणित क्लिटिन में डीएनए दो बार कम होता है, द्विगुणित में . स्पष्ट चयापचय स्थिरता से डीएनए की आनुवंशिक भूमिका का महत्व भी स्पष्ट हो गया था। 1950 के दशक के कोब पर, कई अलग-अलग तथ्य जमा हुए, जिससे पता चला कि अधिकांश उत्परिवर्तजन कारक न्यूक्लिक एसिड के लिए अधिक महत्वपूर्ण हैं, विशेष रूप से डीएनए के लिए (आर। होचकिस, 1949; जी। एफ्रस-टेलर, 1951; ई। फ्रिज़) , 1957 और में।)
न्यूक्लिक एसिड की स्थापित आनुवंशिक भूमिका में विशेष महत्व विभिन्न चरणों और वायरस का थोड़ा प्रजनन है। 1933 में डी. स्लेसिंगर एस्चेरिचिया कोलाई के बैक्टीरियोफेज में डीएनए जानता है। जिस क्षण से डब्ल्यू. स्टेनली (1935, नोबेल पुरस्कार, 1946) ने क्रिस्टलीय अवस्था में टाइयूट्युन मोज़ेक वायरस (TMV) को देखा, प्लांट वायरस में एक नया चरण दिखाई दिया। 1937 - 1938 में आरआर। रोथमस्टेड सिल्स्को पोडारा स्टेशन (इंग्लैंड) के विशेषज्ञ एफ. बोडेन और एन. पिरी ने दिखाया कि उन्होंने बहुत सारे बढ़ते वायरस देखे हैं जो ग्लोब्युलिन नहीं हैं, बल्कि राइबोन्यूक्लियोप्रोटीन हैं और न्यूक्लिक एसिड के बाध्यकारी घटक की तरह काम करते हैं। 1940 के दशक की शुरुआत में, G. Schramm (1940), P. A. Agatov (1941), G. मिलर और W. Stanley (1941) की रचनाएँ प्रकाशित हुईं, जिनमें उन लोगों का उल्लेख किया गया था जिन्हें प्रोटीन घटक के रासायनिक संशोधन का उत्पादन नहीं करना चाहिए। टीएमवी की संक्रामकता से पहले। इसने उन लोगों की ओर इशारा किया कि प्रोटीन घटक वायरस की मंदी की शक्तियों को सहन नहीं कर सकता, क्योंकि वे बहुत सारे सूक्ष्म जीवविज्ञानी का सम्मान करते रहे। वायरस के विकास में न्यूक्लिक एसिड (आरएनए) की आनुवंशिक भूमिका के साक्ष्य पर पुनर्विचार 1956 में वापस ले लिया गया था। तुबिंगन (एफआरएन) के पास जी। श्राम और कैलिफोर्निया (यूएसए) के पास एच। फ्रेनकेल-कोनराथ। उत्तरार्द्ध ने लगभग एक साथ और स्वतंत्र रूप से एक प्रकार के टीएमवी आरएनए को देखा और दिखाया कि यह स्वयं था, न कि प्रोटीन, कम संक्रामक: संक्रमण के परिणामस्वरूप, उनमें ट्युट्युन त्से आरएनए की वृद्धि के परिणामस्वरूप सामान्य वायरल कणों का निर्माण और प्रजनन हुआ। . Tse का मतलब था कि RNA में वायरल प्रोटीन सहित सभी वायरल घटकों के चयन के संश्लेषण के लिए जानकारी होनी चाहिए। 1968 आर. मैं। आर। अताबेकोव ने स्थापित किया है कि प्रोटीन रोजलिन के संक्रमण में एक महत्वपूर्ण भूमिका निभाता है - प्रोटीन की प्रकृति रोजलिन-लॉर्ड्स के स्पेक्ट्रम को निर्धारित करती है।
1957 आर. Frenkel-Konrat ने भंडारण घटकों से TMV के पुनर्निर्माण का बीड़ा उठाया - इस प्रोटीन का RNA। मिश्रित "संकर" से नसों के कई सामान्य हिस्सों को हटा दिया गया था, कुछ आरएनए में यह एक तनाव से था, और प्रोटीन - दूसरे से। इस तरह के संकरों का पतन मुख्य रूप से आरएनए के कारण हुआ था, और वायरस की संतान उस तनाव से संबंधित थी, जिसका आरएनए बाहरी बेमेल कणों से सबसे आम था। ए. गेयरर, जी. शूस्टर और जी. श्राम (1958) और जी. विटमैन (1960 - 1966) के बाद के अध्ययनों से पता चला है कि टीएमवी न्यूक्लिक घटक के रासायनिक संशोधन से इस वायरस में विभिन्न म्यूटेंट की उपस्थिति हो सकती है।
1970 आर. डी। बाल्टीमोर और आर। टेमिन ने स्थापित किया कि आनुवंशिक जानकारी के हस्तांतरण को डीएनए के रूप में आरएनए के रूप में देखा जा सकता है, लेकिन यह भी नहीं। उन्होंने कुछ ऑन्कोजेनिक आरएनए वायरस (ऑनकोर्नवायरस) में एक विशेष एंजाइम, तथाकथित सीरम ट्रांसक्रिपटेस का खुलासा किया है, जो डीएनए को संश्लेषित करने के पूरक आरएनए लांसर्स पर बनाया गया है। इस महान अंतर्दृष्टि ने आरएनए वायरस की आनुवंशिक जानकारी के मास्टर के जीनोम में परिचय के तंत्र को समझना और उनके ऑन्कोजेनिक रोगों की प्रकृति पर नए सिरे से विचार करना संभव बना दिया।
विद्कृत्य न्यूक्लिन अम्ल और उनकी शक्तियों की पुष्टि
न्यूक्लिक एसिड शब्द की शुरुआत जर्मन बायोकेमिस्ट आर. ऑल्टमैन ने 1889 में की थी, उसके बाद इसे अर्ध-जीवन के रूप में 1869 में पेश किया गया था। स्विस चिकित्सक एफ. मिशर। मिशर एकस्ट्रागुव क्लिटिनी मवाद हाइड्रोक्लोरिक एसिड से पतला होता है और कुछ टिज़निव को खींचता है और शुद्ध परमाणु सामग्री की अधिकता को दूर करता है। इस सामग्री को नैदानिक नाभिक के विशिष्ट भाषण द्वारा ध्यान में रखा गया था और इसे योगो न्यूक्लियस कहा जाता था। अपने प्रभुत्व के पीछे, गोरों की उपस्थिति में नाभिक तेजी से कांपते हैं: बेल अधिक खट्टी होती है, सिर्क को नहीं दबाती है, लेकिन नए गुलदस्ते में यह फास्फोरस से भरपूर होता है, यह घास के मैदानों में अच्छा होता है, लेकिन एसिड के सड़ने में नहीं। .
मिशर नाभिक पर अपने प्रेक्षणों के परिणामों को एक जर्नल में प्रकाशन के लिए होप्पे-सीलर को भेजें। उन्होंने एक अनदेखी के फर्श पर बुला के भाषण का वर्णन किया (जैविक फास्फोरस के मामले में, लेकिन केवल लेसिथिन की उपस्थिति में), लेकिन गोपे-ज़ेयलर ने मिशर की टिप्पणियों पर विश्वास नहीं किया, अपनी पांडुलिपि को बदल दिया और अपने स्पाइव्रोबेटनिक को सौंप दिया एम. प्लॉश और एन. हुबाविनु . मिशर के काम "मवाद के रासायनिक गोदाम के बारे में" ने दुनिया को जीवन के दो भाग्य (1871) से भर दिया। उसी समय, होप-सीलर और अन्य स्पाइवोबिटनिक के कार्यों को मवाद, पक्षियों के एरिथ्रोसाइट्स, सांप और अन्य क्लिटिन के गोदाम में प्रकाशित किया गया था। Klіtin और drіzhdzhіv के प्राणियों से दृष्टि के नाभिक के आने वाले तीन भाग्य का प्रोटोज़।
अपने काम में, मिशर ने संकेत दिया कि न्यूक्लिक एसिड की विशिष्टता के विचार को व्यक्त करते हुए, उनके बीच भेद स्थापित करने से पहले विभिन्न न्यूक्लिक एसिड का विस्तृत विस्तार किया जा सकता है। Doslіdzhuyuchi सामन दूध, Mіsher ने स्थापित किया है कि नमक की दृष्टि से उनमें नाभिक पाए जाते हैं और मुख्य प्रोटीन के साथ बंधे होते हैं, जिसे उन्होंने प्रोटामाइन कहा।
1879 में पी. गोप्पे-सीलर की प्रयोगशाला में, ए. कोसेल ने न्यूक्लिक निषेचन का अध्ययन करना शुरू किया। 1881 में पी. मैंने नाभिक में हाइपोक्सैन्थिन देखा है, लेकिन उस समय मैं आधारों के बारे में अधिक संदिग्ध था और यह प्रतिज्ञा कर रहा था कि हाइपोक्सैन्थिन प्रोटीन का क्षरण उत्पाद हो सकता है। 1891 में पी. कोसेल नाभिक के हाइड्रोलिसिस में उत्पादों में, एडेनिन, गुआनिन, फॉस्फोरिक एसिड, और कुक्रू की शक्ति के साथ एक और भाषण का पता चला है। 1910 में न्यूक्लिक एसिड कोसेल के रसायन विज्ञान पर शोध के लिए। नोबेल पुरस्कार से सम्मानित किया गया था।
न्यूक्लिक एसिड की संरचना को समझने में और सफलताएं पी। लेविन और उनके सहयोगियों (1911 - 1934) के अध्ययन से संबंधित हैं। 1911 में पी. लेविन और डब्ल्यू. जैकब्स ने एडीनोसिन और ग्वानोसिन के कार्बोहाइड्रेट घटक की पहचान की; बदबू इसलिए लगाई गई थी ताकि डी-राइबोज इन न्यूक्लियोसाइड के गोदाम में प्रवेश कर सके। 1930 में लेविन ने दिखाया कि डीऑक्सीराइबोन्यूक्लियोसाइड का कार्बोहाइड्रेट घटक 2-डीऑक्सी-डी-राइबोज है। यह ज्ञात हो गया है कि न्यूक्लिक एसिड न्यूक्लियोटाइड द्वारा फॉस्फोराइलेटेड न्यूक्लियोसाइड होने के लिए प्रेरित होते हैं। लेविन ने नोट किया कि न्यूक्लिक एसिड (आरएनए) में मुख्य प्रकार का लिंकेज 2 ", 5" -फॉस्फोडाइस्टर लिंकेज है। त्से उपस्थिति को क्षमा किया गया प्रतीत होता है। अंग्रेजी रसायनज्ञ ए। टॉड (नोबेल पुरस्कार, 1957) और योग वैज्ञानिकों के रोबोटों के साथ-साथ 50 के दशक के कोब पर अंग्रेजी बायोकेमिस्ट आर। मार्खम और जे। स्मिथ के लिए ज़ावद्याकी, यह स्पष्ट हो गया कि लिंकेज का मुख्य प्रकार RNA5 में "- फॉस्फोडाइफिरी लिंक।
लेविन ने दिखाया कि कार्बोहाइड्रेट घटक की प्रकृति से विभिन्न न्यूक्लिक एसिड पर सवाल उठाया जा सकता है: कुछ डीऑक्सीराइबोज पर हमला कर सकते हैं, अन्य - राइबोज। इसके अलावा, दो प्रकार के न्यूक्लिक एसिड को समान आधारों की प्रकृति के अनुसार सौंपा गया था: पेंटोस प्रकार के न्यूक्लिक एसिड में, यूरैसिल का बदला लिया जाता है, और डीऑक्सीपेन्टोज प्रकार के न्यूक्लिक एसिड में थाइमिन होता है। डीऑक्सीपेन्टोज न्यूक्लिक एसिड (आज की शब्दावली में, डीऑक्सीराइबोन्यूक्लिक एसिड - डीएनए) बछड़ों की बड़ी संख्या में थाइमस हड्डियों (क्रोन) में आसानी से देखा जा सकता था। इसके लिए वोन ने थायमोन्यूक्लिक एसिड का नाम ले लिया। न्यूक्लिक एसिड और पेंटोस टाइप (आरएनए) के मूल ने खमीर और गेहूं के कीटाणुओं के प्रमुख रैंक के रूप में कार्य किया। इस प्रकार को अक्सर खमीर न्यूक्लिक एसिड कहा जाता था।
1930 के दशक के सिल पर, एक घटना ने जड़ें जमा लीं, लेकिन बढ़ते क्लिटिन के लिए, एक खमीर-प्रकार का न्यूक्लिक एसिड विशेषता है, और थाइमोन्यूक्लिक एसिड प्राणी क्लिटिन के नाभिक की तुलना में अधिक शक्तिशाली है। दो प्रकार के न्यूक्लिक एसिड - आरएनए और डीएनए - को डेवी और वाइल्ड न्यूक्लिक एसिड भी कहा जाता है। प्रोटीन, जैसा कि ए.एन. बिलोज़र्स्की के शुरुआती अध्ययनों से पता चलता है, न्यूक्लिक एसिड का ऐसा क्षेत्र असत्य है। 1934 में बिलोज़र्स्की ने सबसे पहले पौधों की कोशिकाओं में थाइमोन्यूक्लिक एसिड का पता लगाया: मटर के अंकुरों से, उन्होंने थाइमिन-पाइरीमिडीन बेस को देखा और पहचाना, जो डीएनए की विशेषता है। पोटिम अन्य रोसलिन्स में समय का खुलासा करता है (nasіnі soї, kvasolі)। 1936 में ए एन बिलोज़्स्की और आई। मैं। डबरोव्स्की को किंस्की चेस्टनट के रोपण से प्रारंभिक रूप से डीएनए देखा गया था। इसके अलावा, रोबोटों की एक श्रृंखला, इंग्लैंड में डी. डेविडसन और उनके स्पाइरोबिटनिक द्वारा 40 के दशक में विजयी हुई, ने असंगत रूप से दिखाया कि न्यूक्लिक एसिड (आरएनए) की वृद्धि समृद्ध प्राणियों में पाई जाती है।
डीएनए के लिए साइटोकेमिकल प्रतिक्रिया के आर। फेलगेन और जी। रोसेनबेक (1924) के व्यापक प्रसार और आरएनए के लिए जे। ब्रेचेट (1944) की प्रतिक्रिया ने न्यूक्लिक एसिड के स्थानीयकरण के महत्व के बारे में एक स्पष्ट निष्कर्ष निकालना संभव बना दिया। क्लिटिन ऐसा प्रतीत हुआ कि डीएनए नाभिक में अनुक्रमित होता है, जबकि आरएनए अधिक महत्वपूर्ण रूप से साइटोप्लाज्म में केंद्रित होता है। बाद में पता चला कि आरएनए साइटोप्लाज्म और न्यूक्लियस दोनों में स्थित है, और इसके अलावा, साइटोप्लाज्मिक डीएनए का पता चला था।
जहां तक न्यूक्लिक एसिड की प्राथमिक संरचना के बारे में ज्ञान का सवाल है, 40 के दशक के मध्य तक, पी. लेविन के इस कथन की वैज्ञानिक रूप से पुष्टि की गई थी, जिसके अनुसार न्यूक्लिक एसिड एक प्रकार से प्रेरित थे और टेट्रान्यूक्लियोटाइड के समान शीर्षकों से बनते हैं। ब्लॉक। त्वचीय ब्लॉक में लेविन के विचार के अनुसार, चोटिरी विभिन्न न्यूक्लियोटाइड का बदला लिया जाना चाहिए। न्यूक्लिक एसिड के टेट्रान्यूक्लियोटाइड सिद्धांत ने बायोपॉलिमर विशिष्टता में महत्वपूर्ण योगदान दिया है। यह आश्चर्य की बात नहीं है कि एक जीवित चीज की सभी बारीकियों को केवल प्रोटीन के साथ समझाया गया था, मोनोमर्स की प्रकृति किसी तरह काफी भिन्न होती है (20 अमीनो एसिड)।
थियोरेन्यूक्लियोटाइड और न्यूक्लिक एसिड की पहली सफलता अंग्रेजी रसायनज्ञ जे। गुलैंड (1945 - 1947) के विश्लेषणात्मक डेटा द्वारा बनाई गई थी। जब न्यूक्लिक एसिड की संरचना नाइट्रोजन को सौंपी जाती है, तो नाइट्रोजन के नाइट्रोजन के समतुल्य अनुपात को दूर करना आवश्यक नहीं है, लेकिन लेविन के सिद्धांत का पालन करना पर्याप्त नहीं होगा। न्यूक्लिक एसिड का अवशिष्ट टेट्रान्यूक्लियोटाइड सिद्धांत ई। चारगफ और योग वैज्ञानिकों (1949 - 1951) के परिणामों में आया। उप-आधारों के लिए, जो एसिड हाइड्रोलिसिस के बाद डीएनए से विभाजित होते हैं, कागज पर क्रोमैटोग्राफी द्वारा चारगफ विजयी रहा। त्वचा की त्वचा को स्पेक्ट्रोफोटोमेट्रिक रूप से सटीक रूप से निर्धारित किया गया था। चारगफ ने एक अलग प्रकृति के डीएनए की उपस्थिति में विषुवतीय भिन्नता के महत्व का उल्लेख किया और पहले बताया कि डीएनए प्रजाति विशिष्ट हो सकता है। टिम ने स्वयं जीवित कोशिकाओं में प्रोटीन विशिष्टता की अवधारणा के आधिपत्य को समाप्त किया। विभिन्न दृष्टिकोणों के डीएनए का विश्लेषण करते हुए, चारगफ ने डीएनए संरचना के अनूठे पैटर्न की खोज की और उन्हें तैयार किया, जिसके कारण विज्ञान को चारगफ के नियमों के नाम से जाना गया। इन नियमों के अनुसार, सभी डीएनए के लिए, अंतर से स्वतंत्र, एडेनिन की संख्या थाइमिन (ए = टी) की मात्रा के बराबर होती है, ग्वानिन की मात्रा साइटोसिन (जी = सी) की मात्रा के बराबर होती है। प्यूरीन की मात्रा पाइरीमिडीन + C की मात्रा के बराबर होती है, (G + A = 6-एमिनो समूहों वाले क्षारों की संख्या 6-कीटो समूहों (A+C=G+T) वाले क्षारों की संख्या से अधिक होती है। उन लोगों के बारे में एक ही समय में, इस तरह के सुवोरी kіlkіsnі vіdpovіdnosti के बावजूद, विभिन्न प्रजातियों के डीएनए vіdnoshennia A + T: G + C के मूल्य के लिए vіdrіznyayutsya। कुछ डीएनए में, ग्वानिन और साइटोसिन की मात्रा एडेनिन और थाइमिन की मात्रा से अधिक होती है (चारगाफ को डीएनए जीसी-प्रकार कहा जाता है); अन्य डीएनए ने एडेनिन और थाइमिन का अधिक बदला लिया, कम ग्वानिन और साइटोसिन (क्यूई डीएनए को डीएनए एटी-टाइप कहा जाता था)। डीएनए की संरचना पर ओट्रीमनी चारगफ के डेटा ने आणविक जीव विज्ञान की भूमिका निभाई। बदबू ने ही डीएनए के विकास का आधार बनाया, जिसे 1953 में जे. वाटसन और एफ. क्रिक ने कुचल दिया था।
1938 में शे. डब्ल्यू। एस्टबरी और एफ। बेल ने अतिरिक्त एक्स-रे विवर्तन विश्लेषण का उपयोग करते हुए दिखाया कि डीएनए में आधारों के विमानों को अणु की लंबी धुरी के लंबवत माना जाता है और प्लेटों के द्वि-स्टोस की तरह होता है, जो एक झूठ बोलते हैं एक पर। 1952 - 1953 तक दुनिया ने एक्स-रे विवर्तन विश्लेषण की तकनीक को सिद्ध किया है। संचित vіdomostі, scho ने दर्जनों okremih zv'yazkіv ta kutakh nahily के बारे में न्याय करने की अनुमति दी। इसने डीएनए अणु के शुगर-फॉस्फेट बैकबोन में पेंटोस अवशेषों के उन्मुखीकरण की प्रकृति को सबसे बड़ी स्पष्टता के साथ प्रकट करना संभव बना दिया। 1952 आर. एस. फारबर्ग ने डीएनए के दो समझदार मॉडल प्रस्तावित किए, उन्होंने एक एकल-फंसे हुए अणु की कल्पना की, जो अपने आप मुड़ा या मुड़ा हुआ था। बुडोव के डीएनए बुला का कोई कम सट्टा मॉडल 1953 में प्रस्तावित नहीं किया गया था। एल. पॉलिंग (नोबेल पुरस्कार विजेता, 1954) और आर. कोरी। इस मॉडल में, तीन मुड़ डीएनए लेंसों ने एक लंबा हेलिक्स बनाया, जिसके कतरन को फॉस्फेट समूहों द्वारा दर्शाया गया था, और सिलवटों को नए लोगों द्वारा बदल दिया गया था। 1953 तक, एम. विल्किंस और आर. फ्रैंकलिन ने डीएनए के अधिक सटीक एक्स-रे पैटर्न लिए। वर्तमान विश्लेषण ने फारबर्ग, पॉलिंग और कोरी के मॉडलों की पूर्ण असंभवता को दिखाया। 1953 में चार मोनोमर्स के विभिन्न आणविक मॉडल और एक्स-रे विवर्तन विश्लेषण के डेटा, जे। वाटसन और एफ। क्रिक को बताते हुए, चारगफ का विकोरिस्ट डेटा। dіyshli vysnovka, scho DNA अणु एक डबल-स्ट्रैंडेड हेलिक्स हो सकता है। चारगफ के नियमों ने डीएनए मॉडल के प्रस्ताव के लिए संभावित आदेश देने वाले ठिकानों की संख्या को तेजी से पीछे छोड़ दिया; बदबू ने वाटसन और क्रिक को सुझाव दिया कि डीएनए अणुओं में आधारों की एक विशिष्ट जोड़ी हो सकती है - थाइमिन के साथ एडेनिन, और साइटोसिन के साथ ग्वानिन। दूसरे शब्दों में, डीएनए की एक भाषा में एडेनिन हमेशा दूसरे लांस में थाइमिन दिखाता है, और डीएनए के उसी लालटेन में गुआनिन दूसरे में साइटोसिन दिखाता है। टिम वाटसन और क्रिक ने स्वयं पहले पूरक डीएनए के सिद्धांत के विनीत महत्व को तैयार किया, जिसका अर्थ है कि डीएनए का एक लैंसेट दूसरे का पूरक है, ताकि एक लैंसेट की घटना का क्रम स्पष्ट रूप से दूसरे (पूरक) लैंसेट में होने वाली घटनाओं के अनुक्रम को इंगित करता है। . यह स्पष्ट हो गया कि डीएनए की संरचना में सटीक निर्माण की क्षमता है। डीएनए के अस्तित्व का त्स्या मॉडल nі zagaloviznanoyu। 1962 में क्रिक, वाटसन और विल्किंस को डीएनए की संरचना को समझने के लिए। नोबेल पुरस्कार से सम्मानित किया गया था।
यह ध्यान दिया जाना चाहिए कि मैक्रोमोलेक्यूल्स के सटीक निर्माण और क्षय सूचना के हस्तांतरण के लिए एक तंत्र का विचार हमारे क्षेत्र में उत्पन्न हुआ। 1927 आर. एन.के. कोल्टसोव ने एक बयान दिया है, कि कोशिकाओं के प्रजनन के दौरान, स्पष्ट मूल अणुओं के सटीक ऑटोकैटलिटिक प्रजनन की तर्ज पर अणुओं का प्रजनन देखा जाता है। सच है, भले ही कोल्टसोव ने डीएनए अणुओं को नहीं, बल्कि प्रोटीन प्रकृति के अणुओं को शक्ति दी, उनमें से किसी के कार्यात्मक महत्व के बारे में कुछ भी नहीं पता था। मैक्रोमोलेक्यूल्स के ऑटोकैटलिटिक निर्माण और आवर्ती शक्तियों के हस्तांतरण के तंत्र के बारे में बहुत ही विचार एक भविष्यवक्ता निकला: यह आधुनिक आणविक जीव विज्ञान का एक मौलिक विचार बन गया।
A. N. Bilozersky A. S. Spirin, G. N. Zaitseva, B. F. Vanyushin, S. O. Urison, A. S. Antonov की प्रयोगशाला में आयोजित विभिन्न जीवों ने चारगफ द्वारा प्रकट नियमितताओं की पुष्टि की, साथ ही वाटसन और क्रिक द्वारा सुझाए गए डीएनए के अस्तित्व के आणविक मॉडल के साथ समानता की पुष्टि की। . कई अध्ययनों से पता चला है कि विभिन्न बैक्टीरिया, कवक, शैवाल, एक्टिनोमाइसेट्स, शैवाल, स्पिनलेस और स्पिनलेस के डीएनए गोदाम के लिए विशिष्ट हो सकते हैं। गोदाम में विशेष रूप से तेज (आधारों के एटी-जोड़े में परिवर्तन) सूक्ष्मजीव में अभिव्यक्तियाँ, एक महत्वपूर्ण टैक्सोनॉमिक संकेत दिखाती हैं। आधुनिक पौधों और जीवों में, डीएनए गोदाम में बदलाव काफी कमजोर हैं। Alce zovsim का मतलब यह नहीं है कि उनका डीएनए कम विशिष्ट है। क्रिमियम गोदाम, स्थापित विशिष्टता के साथ, डीएनए लेंस में इसके अनुक्रम को महत्वपूर्ण रूप से सौंपा गया है।
डीएनए और आरएनए के गोदाम में सबसे महत्वपूर्ण ठिकानों का क्रम एडिटिव नाइट्रोजनस बेस के रूप में सामने आया। इस प्रकार, जी। व्हाइट (1950) बढ़ते जीवों के डीएनए गोदाम में 5-मिथाइलसिटोसिन को जानता था, और डी। डन और जे। स्मिथ (1958) ने कुछ डीएनए में एडेनिन मिथाइलेशन का पता लगाया। लंबे समय तक, मिथाइलसिटोसिन को जीवित जीवों की आनुवंशिक सामग्री के विशिष्ट चावल में पेश किया गया था। 1968 आर. A. N. Bilozersky, B. F. Vanyushin और N. A. Kokurina ने स्थापित किया कि नसों को बैक्टीरिया के डीएनए में भी दर्ज किया जा सकता है।
1964 में एम। गोल्ड और जे। हर्विट्ज़ ने एंजाइमों के एक नए वर्ग की खोज की जो डीएनए के प्राकृतिक संशोधन को प्रभावित करते हैं - मिथाइलेशन। इस जांच के बाद, यह स्पष्ट हो गया कि नाबालिगों (जो कम संख्या में पाए जाते हैं) को विशेष अनुक्रमों में अतिरिक्त साइटोसिन और एडेनिन के विशिष्ट मिथाइलेशन के परिणामस्वरूप तैयार डीएनए पोलीन्यूक्लियोटाइड लांस पर पहले से ही दोषी ठहराया जाता है। ज़ोक्रेमा, बी. एफ. वनुशिन की श्रद्धांजलि के लिए, हां. आई. Bur'yanova और A. N. Bilozerskogo (1969) Escherichia coli के डीएनए में एडेनिन का मिथाइलेशन कोडन को समाप्त करने में पाया जा सकता है। A. N. Bilozersky और spivrobitnikov (1968 - 1970), साथ ही M. Meselson (USA) और V. Arber (स्विट्जरलैंड) (1965 - 1969) के अनुसार, मिथाइलेशन डीएनए अणुओं को तह तंत्र के विशिष्ट न्यूक्लियस भाग में एक अनूठा पैटर्न देता है, जो क्लिटिन में डीएनए संश्लेषण को नियंत्रित करता है। दूसरे शब्दों में, इस डीएनए के मिथाइलेशन की प्रकृति उन लोगों के लिए पोषण को इंगित करती है जो इस क्लिटिन में गुणा कर सकते हैं।
लगभग उसी समय, डीएनए मिथाइलिस और प्रतिबंध एंडोन्यूक्लाइज के उस गहन टीकाकरण को देखा गया था; 1969 - 1975 पीपी। इन एंजाइमों द्वारा डीएनए में पेश किए गए न्यूक्लियोटाइड अनुक्रमों को डाला गया था (एक्स बॉयर, एक्स स्मिथ, एस लिन, के। मरे)। विभिन्न डीएनए के हाइड्रोलिसिस में, प्रतिबंध एंजाइम को एक ही "चिपचिपा" सिरों के साथ बड़े टुकड़ों को पूरा करने की अनुमति है। यह जीन की संरचना का विश्लेषण करने की संभावना देता है, क्योंकि यह छोटे वायरस (डी। नाथन, जेड एडलर, 1973 - 1975) में टूट जाता है, और विभिन्न जीनोम का निर्माण करता है। इन विशिष्ट प्रतिबंध एंजाइमों की मान्यता के साथ, आनुवंशिक इंजीनियरिंग एक स्पष्ट वास्तविकता बन गई है। छोटे प्लास्मिड जीन के मामले में, विभिन्न प्रकार के जीन के डीएनए को विभिन्न प्रकार की कोशिकाओं में इंजेक्ट करना पहले से ही आसान है। इसलिए, एक नए प्रकार के जैविक रूप से सक्रिय प्लास्मिड को अपनाया गया, जो डेकी एंटीबायोटिक्स (एस। कोएन, 1973) को स्थिरता देता है, एस्चेरिचिया कोलाई के प्लास्मिड में टॉड और ड्रोसोफिला के राइबोसोमल जीन की शुरूआत (जे। मोरो, 1974; एक्स बॉयर, डी। हॉगनेस, आर।, 1974 - 1975)। इस प्रकार, हमने मौलिक रूप से नए जीवों को विभिन्न जीनों के जीन पूल में पेश करके उनके चयन के लिए वास्तविक रास्ते खोले हैं। Tse vіdkrittya को लोगों के लाभ के लिए सीधा किया जा सकता है।
1952 आर. जी। व्हाइट और एस। कोहेन ने दिखाया कि टी-जोड़ी चरणों के डीएनए में एक अज्ञात आधार है - 5-हाइड्रॉक्सीमिथाइलसिटोसिन। ई। वोल्किन और आर। सिनशाइमर (1954) और कोहेन (1956) के काम से यह स्पष्ट हो गया कि अतिरिक्त ऑक्सीमिथाइलसिटोसिन आंशिक रूप से या आंशिक रूप से ग्लूकोसिडेट हो सकता है, जिसके बाद फेज डीएनए अणु हाइड्रोलाइटिक न्यूक्लीज से चोरी होता प्रतीत होता है।
1950 के दशक के कोब पर, डी। डन और जे। स्मिथ (इंग्लैंड), एस। ज़मेनहोफ़ (यूएसए) और ए। वेकर (एफआरएन) का काम स्पष्ट हो गया कि डीएनए में इनोड्स की जगह, बेस के बहुत सारे टुकड़े एनालॉग शामिल हो सकते हैं। 50% थाइम तक। एक नियम के रूप में, क्यूई प्रतिस्थापन को प्रतिकृति, डीएनए के प्रतिलेखन और म्यूटेंट की उपस्थिति तक अनुवाद के दौरान क्षमा के लिए लाया जाता है। इस प्रकार, जे। मर्मुर (1962) ने स्थापित किया कि कुछ चरणों के डीएनए थाइमिन को ऑक्सीमिथाइलुरैसिल से बदल देते हैं। 1963 पी. मैं। ताकाहाशी और जे। मर्मुर ने दिखाया कि एक फेज के डीएनए में यूरैसिल थाइमिन की जगह लेता है। इस रैंक में, एक और सिद्धांत कहा जाता था, जिसके बाद न्यूक्लिक एसिड पहले जोड़ा गया था। पी. लेविन के काम के दौरान, यह महत्वपूर्ण था कि थाइमिन डीएनए की एक विशेषता है, और यूरैसिल आरएनए की एक विशेषता है। यह स्पष्ट हो गया कि यह संकेत आवश्यक रूप से सतही नहीं है, और दो प्रकार के न्यूक्लिक एसिड की रासायनिक प्रकृति का महत्वपूर्ण विचार, जैसा कि आज लगता है, केवल कार्बोहाइड्रेट घटक की प्रकृति है।
फेज कल्चर के समय में, न्यूक्लिक एसिड के संगठन के बहुत सारे महत्वहीन संकेत सामने आए। जेड 1953 यह महत्वपूर्ण था कि डीएनए दो-फंसे रैखिक अणु है, और आरएनए एकल-फंसे से कम है। 1961 में साइट का अपहरण कर लिया गया था जब आर। सिनशाइमर ने दिखाया कि फेज X 174 के डीएनए को एकल-फंसे हुए रिंग अणु द्वारा दर्शाया गया है। तब यह सच था कि ऐसे रूप में डीएनए केवल वनस्पति फेज कण में मौजूद होता है, और इस फेज के डीएनए का प्रतिकृति रूप भी दो-फंसे होता है। इसके अलावा, यह अविश्वसनीय रूप से स्पष्ट था कि कुछ वायरस के आरएनए दो-फंसे हो सकते हैं। 1962 में आरएनए के एक नए प्रकार के मैक्रोमोलेक्यूलर संगठन की खोज की गई थी। पी। गोमेटोस, आई। कुछ वायरस जानवरों में टैम और अन्य उत्तराधिकारी और रोसलिन की प्रारंभिक सूजन के वायरस में। हाल ही में वी.आई. एगोल और ए.ए. बोगदानोव (1970) ने स्थापित किया कि रैखिक आरएनए अणुओं की क्रीम भी बंद या चक्रीय अणु है। उनके द्वारा चक्रीय डबल-स्ट्रैंडेड आरएनए का पता लगाया गया था, जोक्रेमा, एन्सेफेलोमाइलोकार्डिटिस वायरस में। Zavdyaki रोबोट X. Deveaux, L. Tinoko, T. I. तिखोनेंको, ई. आई. बुडोव्स्की और अन्य (1960 - 1974) बैक्टीरियोफेज में आनुवंशिक सामग्री के संगठन (तह) के मुख्य प्रकार बन गए।
उदाहरण के लिए, 1950 के अमेरिकी अध्ययन, पी. डॉट ने स्थापित किया है कि डीएनए विकृतीकरण हीटिंग के दौरान होता है, जो आधारों के जोड़े के बीच पानी के लिंक के विकास और पूरक लेंस के भेदभाव के साथ होता है। यह प्रक्रिया "सर्पिल-कॉइल" क्रिस्टल के लिए एक चरण संक्रमण की प्रकृति में है और माना जाता है कि क्रिस्टल पिघल जाते हैं। इसलिए, डीएनए डॉटी के थर्मल विकृतीकरण की प्रक्रिया को डीएनए मेल्टिंग कहा जाता है। उचित शीतलन के साथ, अणुओं का पुनर्विकास होता है, जिससे पूरक हिस्सों का पुनर्जन्म होता है।
1960 में पुनर्जीवन का सिद्धांत विभिन्न सूक्ष्मजीवों के डीएनए के "संकरण" के निर्दिष्ट चरण के लिए जे। मर्मुर और के। शिल्डक्राट। वैसे, ई. बोल्टन और बी. मैकार्थी ने तथाकथित डीएनए-एगर कॉलम पद्धति की शुरुआत करके इस दृष्टिकोण को सिद्ध किया। विभिन्न डीएनए के न्यूक्लियोटाइड अनुक्रम के समरूपता स्तर के प्रजनन और विभिन्न जीवों की आनुवंशिक स्पोरिडिटी के निर्धारण के लिए यह विधि अपरिहार्य साबित हुई। जे. मंडेल और ए. हर्षे द्वारा वर्णित विधि में डीएनए का विद्कृत दोती विकृतीकरण * (1960) मिथाइलेटेड एल्ब्यूमिन पर क्रोमैटोग्राफी और ग्रेडिएंट्स पर सेंट्रीफ्यूजेशन (1957 में एम। मेसेलसन, एफ। स्टाल और डी। विनोग्राद द्वारा पृथक्करण की विधि) पृथक्करण , चार पूरक डीएनए लेंसों का देखा और विश्लेषण। उदाहरण के लिए, वी। स्ज़ीबाल्स्की (यूएसए), विकोरी का उपयोग लैम्ब्डा फेज डीएनए के लिए किया गया था, जो 1967 - 1969 पीपी। रहुवती (एस। स्पाइजेलमैन, 1961) में दिखा। यह ध्यान दिया जाना चाहिए कि लैम्ब्डा फेज के डीएनए के दोनों लेंसों के आनुवंशिक महत्व के बारे में विचार सबसे पहले एसआरएसआर एस में विकसित किया गया था। ब्रेस्लर (1961)।
* (बैक्टीरिया और वायरस के आनुवंशिकी पर काम करने के लिए, ए। हर्षे को एम। डेलब्रुक और एस। लुरिया के साथ संयुक्त रूप से 1969 से सम्मानित किया गया। नोबेल पुरुस्कार।)
जीनोम के संगठन और कार्यात्मक गतिविधि को समझने के लिए, डीएनए के न्यूक्लियोटाइड अनुक्रम का असाइनमेंट सर्वोपरि है। इस तरह के पदनाम के तरीकों की खोज दुनिया की समृद्ध प्रयोगशालाओं में की जाती है। 1950 के दशक की शुरुआत से एम. बीयर और उनके सहयोगियों ने इलेक्ट्रॉन माइक्रोस्कोपी की मदद से डीएनए अनुक्रम स्थापित करने का प्रयास किया, लेकिन अभी तक सफलता नहीं मिली। 1950 के दशक के कोब पर, सिनशाइमर, चारगफ के पहले कार्यों और डीएनए के एंजाइमेटिक क्षरण के अन्य हालिया अध्ययनों से, यह स्पष्ट हो गया कि वितरण के डीएनए अणु में न्यूक्लियोटाइड्स में अंतर अराजक नहीं है, बल्कि असमान है। अंग्रेजी रसायनज्ञ सी. बार्टन (1961) के आंकड़ों के अनुसार, पाइरीमिडीन (लगभग 70%) ब्लॉकों की तरह दिखने में अधिक महत्वपूर्ण है। ए.एल. माज़िन और बी.एफ. वनुशिन (1968 - 1969) ने स्थापित किया कि विभिन्न डीएनए स्तर पाइरीमिडीन रुकावट के विभिन्न चरणों को जन्म दे सकते हैं और जीवों के जीवों के डीएनए में, दुनिया में निचले से उच्चतर में संक्रमण में उल्लेखनीय वृद्धि हुई है। साथ ही जीवों का विकास उनके जीनोम की संरचना में भी देखा जाता है। इसी कारण से, विकासवादी प्रक्रिया को समझने के लिए, सामान्य तौर पर, न्यूक्लिक एसिड की संरचना के विकास का विशेष महत्व है। जैविक रूप से महत्वपूर्ण पॉलिमर और डीएनए की संरचना का विश्लेषण फ़ाइलोजेनेटिक्स और टैक्सोनॉमी के समृद्ध निजी पोषण के विकास के लिए अत्यंत महत्वपूर्ण है।
यह स्पष्ट है कि अंग्रेजी शरीर विज्ञानी ई। लैंकेस्टर, जिन्होंने मोलस्क के हीमोग्लोबिन को विकसित किया, जिन्होंने ठीक 100 वर्षों तक आणविक जीव विज्ञान के विचारों को व्यक्त करते हुए लिखा: महत्वपूर्ण अर्थ z'yasuvannya के लिए isstorії pojzhennya, जैसे कि यह razbіzhnostі में है। हम आणविक संगठन और जीवों के कामकाज में स्पष्ट रूप से पहचान स्थापित कर सकते हैं, हम विभिन्न जीवों के समान विकास में और अधिक तेज़ी से विकसित हो सकते हैं, रूपात्मक संकेतों के आधार पर कम। , स्को "सभी रूपात्मक विशेषताओं के आधार पर, इस तरह के वर्गीकरण के आधार पर, हम देख सकते हैं, बहुत ही जैव रासायनिक विशेषताओं को झूठ बोलना" **।
* (ईआर लैंकेस्टर। उबेर दास वोरकॉमन वॉन हीमोग्लोबिन इन डेन मस्केलन डेर मोलस्केन अंड डाई वर्ब्रिटुंग डेसेलबेन इन डेन लेबेन्डिजेन ऑर्गेनिस्मेन।- "पफ्लुगर" का आर्किव फर डाई गेसम्टे फिजियोल।, 1871, बीडी 4, 319।)
** (वी एल कोमारोव। चयनित कार्य, वी. 1. एम.-एल., एसआरएसआर की विज्ञान अकादमी का प्रकार, 1945, पृष्ठ 331।)
ए.वी. ब्लागोविशचेन्स्की और एस.एल. इवानोव ने 1920 के दशक में अपने जैव रासायनिक संरचना के कालानुक्रमिक विश्लेषण के आधार पर जीवों के सिस्टमैटिक्स के वर्तमान विकास की खोज के लिए हमारे देश में पहला कदम उठाया (div। ch। 2)। टैक्सोनोमिस्ट्स के लिए प्रोटीन और न्यूक्लिक एसिड की संरचना का पंक्ति विश्लेषण अधिक से अधिक उपयोगी होता जा रहा है (डिव। सेक्शन 21)। आणविक जीव विज्ञान की यह पद्धति प्रणाली में कई प्रजातियों की स्थापना को स्पष्ट करना संभव बनाती है, और जीवों के वर्गीकरण के सिद्धांतों पर आश्चर्यचकित करने के लिए एक नए तरीके से, जो कभी-कभी पूरी प्रणाली को धुंध में देखते हैं, जैसे यह बन गया है, उदाहरण के लिए, सूक्ष्मजीव के सिस्टमैटिक्स से। निस्संदेह, जीनोम की संरचना के भविष्य के विश्लेषण में, यह केमोसिस्टमैटिक जीवों में केंद्रीय स्थान पर कब्जा कर लेगा।
आणविक जीव विज्ञान के विकास के लिए बहुत महत्व डीएनए प्रतिकृति और प्रतिलेखन के तंत्र की कम समझ है (डिव। खंड 24)।
प्रोटीन जैवसंश्लेषण
प्रोटीन जैवसंश्लेषण की वायरल समस्या में एक महत्वपूर्ण व्यवधान न्यूक्लिक एसिड के विकास में सफलता से जुड़ा है। 1941 पी. टी. कास्पर्सन (स्वीडन) और 1942 में पैदा हुए जे। ब्रेचेट (बेल्जियम) ने उन लोगों पर ध्यान दिया जिन्होंने सक्रिय प्रोटीन संश्लेषण के साथ ऊतकों में आरएनए की मात्रा में वृद्धि की है। बदबू दीशली विस्नोव्का रही है, स्को राइबोन्यूक्लिक एसिड प्रोटीन संश्लेषण की प्राथमिक भूमिका निभाते हैं। 1953 आर. दूसरी ओर, ई। गेल और डी। फॉक्स ने प्रोटीन जैवसंश्लेषण में आरएनए की अप्रत्यक्ष भागीदारी के प्रत्यक्ष प्रमाण को छीन लिया: इन कारणों से, राइबोन्यूक्लिज़ ने बैक्टीरियल क्लिटिन के लाइसेट्स में अमीनो एसिड के समावेश को महत्वपूर्ण रूप से दबा दिया। वी. ओल्फ़्री, एम. डेली और ए. मिर्स्की (1953) द्वारा लीवर होमोजेनेट्स पर अनुरूप डेटा लिया गया था। पिज़्निशे येगे। गेल, दूध देने के बाद प्रोटीन संश्लेषण में आरएनए की भूमिका के बारे में एक सही विचार के साथ आए, कि सेलुलर सिस्टम में प्रोटीन संश्लेषण की सक्रियता अप्राकृतिक प्रकृति के एक अन्य भाषण के प्रभाव में थी। 1954 पी. पी। ज़मिटनिक, डी। लिटिलफ़ील्ड, आर। बी। हेसिन-लुरेट और अन्य ने दिखाया कि अमीनो एसिड का सबसे सक्रिय समावेश उप-कोशिकीय कणों के समृद्ध आरएनए अंशों में पाया जाता है - माइक्रोसोम। पी। ज़मेचनिक और ई। केलर (1953 - 1954) ने दिखाया कि एटीपी पुनर्जनन के मस्तिष्क में सतह पर तैरनेवाला अंश की उपस्थिति में अमीनो एसिड का समावेश महत्वपूर्ण रूप से देखा गया था। P. Sikevitz (1952) और M. Hoagland (1956) ने सतह पर तैरनेवाला रिनडीन से एक प्रोटीन अंश (pH 5 अंश) देखा, जो माइक्रोसोम में अमीनो एसिड को शामिल करने के लिए अत्यधिक उत्तेजक था। सतह पर तैरनेवाला घर में कई प्रोटीन कम आणविक भार आरएनए के विशेष वर्ग पाए गए, जिन्हें अब परिवहन आरएनए (टीआरएनए) कहा जाता है। 1958 पी. होगलैंड और पोमिचनिक, पी। बर्ग, आर। स्वीट, और एफ। एलन और अन्य ने दिखाया है कि त्वचीय अमीनो एसिड के सक्रियण के लिए अपने स्वयं के विशेष एंजाइम, एटीपी और विशिष्ट टीआरएनए की आवश्यकता होती है। यह स्पष्ट हो गया कि tRNA एडेप्टर की महत्वपूर्ण भूमिका निभाते हैं, ताकि वे, जैसा कि वे जानते हैं, बनने वाले प्रोटीन अणु में आवश्यक अमीनो एसिड के न्यूक्लिक मैट्रिक्स (iRNA) से जुड़ जाते हैं। इन अध्ययनों ने एफ. क्रिक (1957) की एडेप्टर परिकल्पना की पुष्टि की, जिसने कोशिकाओं में पॉलीन्यूक्लियोटाइड एडेप्टर के उपयोग को स्थानांतरित कर दिया, जो न्यूक्लिक मैट्रिक्स पर अमीनो एसिड अतिरिक्त प्रोटीन के सही आसवन के लिए आवश्यक हैं, जो संश्लेषित होते हैं। संयुक्त राज्य अमेरिका में एफ. लिपमैन (नोबेल पुरस्कार, 1953) की प्रयोगशाला में एफ. चैपविले (1962) की समृद्ध फ्रांसीसी शिक्षाओं ने पहले ही गर्मजोशी से और स्पष्ट रूप से दिखाया कि प्रोटीन अणु में अमीनो एसिड का विशेषीकरण आ गया है। क्रिक की अनुकूली परिकल्पना को होगलैंड और पोमिचनिक के रोबोटों में सही ठहराया गया था।
1958 तक, प्रोटीन संश्लेषण के निम्नलिखित मुख्य चरण घर बन गए: 1) एक विशिष्ट एंजाइम द्वारा "पीएच 5 अंश" के साथ अमीनो एसिड की सक्रियता एटीपी की उपस्थिति में एमिनोएसिलेडेनिलेट के समाधान के साथ; 2) एडीनोसिन मोनोफॉस्फेट (एएमपी) से विशिष्ट टीआरएनए में सक्रिय अमीनो एसिड के अलावा; 3) माइक्रोसोम के साथ एमिनोएसिल-टीआरएनए (एमिनो एसिड के साथ टीआरएनए नेवांटेज) का लिंकेज और व्यवहार्य टीआरएनए के साथ प्रोटीन में अमीनो एसिड का समावेश। होगलैंड (1958) ने संकेत दिया कि प्रोटीन संश्लेषण के अंतिम चरण में ग्वानोसिन ट्राइफॉस्फेट (जीटीपी) की आवश्यकता होती है।
परिवहन आरएनए और जीन संश्लेषण
टीआरएनए का पता लगाने के बाद, न्यूक्लियोटाइड अनुक्रम के विभाजन और असाइनमेंट के लिए सक्रिय खोज शुरू हुई। अमेरिकी बायोकेमिस्ट आर होली की सबसे बड़ी सफलता। 1965 में खमीर से ऐलेनिन टीआरएनए की संरचना स्थापित करने के बाद। राइबोन्यूक्लिअस (ग्वानील आरएनए-एएसई और अग्नाशयी आरएनए-एएस) की मदद के लिए, हाले ने न्यूक्लिक एसिड अणु को टुकड़ों में विभाजित किया, त्वचा में न्यूक्लियोटाइड अनुक्रम सौंपा और फिर पूरे एलेनिन टीआरएनए अणु के अनुक्रम का पुनर्निर्माण किया। न्यूक्लियोटाइड अनुक्रम के विश्लेषण का उद्देश्य ब्लॉक विधि का नाम हटाना था। होली की योग्यता को आरएनए अणु को छोटे टुकड़ों में, यहां तक कि बड़े पैमाने पर, और यहां तक कि बड़े टुकड़ों (क्वार्टर और आधा) में विभाजित करने का तरीका सीखने का मुख्य कारण माना जाता था। इससे छोटे टुकड़ों को एक साथ सही ढंग से इकट्ठा करना और पूरे टीआरएनए अणु (नोबेल पुरस्कार, 1968) का एक ही न्यूक्लियोटाइड अनुक्रम बनाना संभव हो गया।
दुनिया की समृद्ध प्रयोगशालाओं में ozbroєnnya पर Tsej priyom vіdrazu buv priynyaty। एसआरएसआर और वहां में अगले दो वर्षों के दौरान, किल्कोह टीआरएनए के क्रॉस-सेक्शन की प्राथमिक संरचना को समझ लिया गया। ए.ए. बेव (1967) और शोधकर्ताओं ने सबसे पहले यीस्ट वेलिन टीआरएनए में न्यूक्लियोटाइड के अनुक्रम की स्थापना की। अब तक, एक दर्जन से अधिक अलग-अलग टीआरएनए पहले ही पैदा हो चुके हैं। एक निर्दिष्ट न्यूक्लियोटाइड अनुक्रम के लिए एक अनूठा रिकॉर्ड कैम्ब्रिज में एफ। सेंगर और जी। ब्राउनली द्वारा स्थापित किया गया था। निम्नलिखित शोधकर्ताओं ने सब-ऑलिगोन्यूक्लियोटाइड्स की विधि को पूरी तरह से विकसित किया और कोलीफॉर्म कोशिकाओं (1968) से तथाकथित 5 एस (राइबोसोमल) आरएनए के अनुक्रम की स्थापना की। Qia RNA 120 न्यूक्लियोटाइड सरप्लस से बना है और, tRNA के आधार पर, अतिरिक्त छोटे आधारों का विरोध नहीं करता है, इस प्रकार न्यूक्लियोटाइड अनुक्रम का विश्लेषण करना आसान बनाता है, अणु के कई टुकड़ों के लिए अद्वितीय स्थलों के रूप में कार्य करता है। दिन के इस समय में, जे। एबेल (फ्रांस) और अन्य हाल के लोगों की प्रयोगशाला में पुराने राइबोसोमल आरएनए और अन्य वायरल आरएनए के रोबोट अनुक्रम द्वारा सेंगर और ब्राउनल विधि का सफलतापूर्वक परीक्षण किया गया है।
ए.ए. बेव और शोधकर्ताओं (1967) ने दिखाया कि विस्तारित नेवीपिल वेलिन टीआरएनए प्राथमिक संरचना में दोष की परवाह किए बिना अपनी मैक्रोमोलेक्यूलर संरचना को एक अलग तरीके से बनाए रखता है, लेकिन बाहरी (देशी) अणु की एक कार्यात्मक गतिविधि हो सकती है। बाद वाला पिदखिद - गीत के टुकड़ों को हटाने के बाद एक कटे हुए मैक्रोमोलेक्यूल का पुनर्निर्माण - और भी अधिक आशाजनक प्रतीत होता है। एक ही समय में व्यापक रूप से vikoristovuєtsya z'yasuvannya okremih dіlyanok शांत ची इन्शिह tRNA की कार्यात्मक भूमिका।
शेष विश्व में, व्यक्तिगत tRNA की क्रिस्टल-आधारित तैयारियों के विकास में बड़ी सफलता प्राप्त हुई है। उसी समय, संयुक्त राज्य अमेरिका और इंग्लैंड में कई प्रयोगशालाओं में, और भी समृद्ध tRNA को क्रिस्टलीकृत करना संभव था। इससे मुझे अतिरिक्त एक्स-रे विवर्तन विश्लेषण के लिए tRNA की संरचना निर्धारित करने का अवसर मिला। 1970 आर. आर बॉक ने विस्कॉन्सिन विश्वविद्यालय द्वारा बनाए गए कई टीआरएनए के पहले रेडियोग्राफ और ट्रिविमर मॉडल प्रस्तुत किए। यह मॉडल टीआरएनए में चार कार्यात्मक रूप से सक्रिय कोशिकाओं के स्थानीयकरण की पहचान करने और इन अणुओं के कामकाज के मुख्य अवरोध को समझने में मदद करता है।
प्रोटीन संश्लेषण के तंत्र को प्रकट करने के लिए सबसे महत्वपूर्ण मूल्य और इस प्रक्रिया की विशिष्टता की समस्या आनुवंशिक कोड की प्रकृति को समझने के लिए पर्याप्त नहीं है (डिव। खंड 24), लेकिन अतिशयोक्ति के बिना यह विचार करना संभव है कि कैसे विजय की विजय XX सदी का प्राकृतिक विज्ञान।
आर. होली द्वारा टीआरएनए की प्राथमिक संरचना की खोज ने जी. कोरनी* (यूएसए) को ऑलिगोन्यूक्लियोटाइड्स के संश्लेषण के लिए एक रास्ता दिया और उन्हें एक एकल जैविक संरचना के संश्लेषण के लिए निर्देशित किया - एक डीएनए अणु जो एलेनिन टीआरएनए को एन्कोड करता है। कुरान के पूरा होने से 15 साल पहले शॉर्ट ऑलिगोन्यूक्लियोटाइड्स का रासायनिक संश्लेषण 1970 में पूरा हुआ था। पहला, zdіysnenim जीन संश्लेषण। 8-12 न्यूक्लियोटाइड्स की कुरान और योग वर्तनी को रासायनिक विधि द्वारा 8-12 न्यूक्लियोटाइड पुर्जों के ज़ावोडोवका के छोटे टुकड़ों द्वारा संश्लेषित किया गया था। किसी दिए गए न्यूक्लियोटाइड अनुक्रम से Cy टुकड़े अनायास ही 4-5 न्यूक्लियोटाइड के ओवरलैप के साथ दो-फंसे पूरक किस्में बनाते हैं। बाद में, डीएनए-लिगेज एंजाइम की मदद से शमतकी को आवश्यक क्रम में, क्रमिक रूप से, अंत से अंत तक पकाने के लिए। इस तरह, डीएनए अणुओं की प्रतिकृति को निर्देशित करने के लिए, ए कोर्नबर्ग ** (डिव। सेक्शन 24) के बाद, कुरान ने प्राकृतिक डबल-स्ट्रैंडेड डीएनए के एक अणु को फिर से बनाने के लिए डिज़ाइन किए गए प्रोग्राम की पृष्ठभूमि के पीछे बनाया। हाले द्वारा वर्णित टीआरएनए अनुक्रम। संक्रमण का एक समान क्रम अन्य जीनों के संश्लेषण के लिए किया जाता है (एम। एम। कोलोसोव, जेड। ए। शबरोवा, डी। जी। नॉररे, 1970 - 1975)।
* (जी. कुरान और एम. निरेनबर्ग के आनुवंशिक कोड को पूरा करने के लिए 1968 में सम्मानित किया गया था। नोबेल पुरुस्कार।)
** (ए. कोर्नबर्ग द्वारा पोलीमरेज़ डीएनए संश्लेषण की मान्यता के लिए, और 1959 में एस. ओचोआ द्वारा आरएनए के संश्लेषण के लिए। नोबेल पुरस्कार से सम्मानित किया गया था।)
माइक्रोसोम, राइबोसोम, अनुवाद
1950 के दशक में, यह महत्वपूर्ण हो गया कि क्लिटिन में प्रोटीन संश्लेषण का केंद्र माइक्रोसोम था। माइक्रोसोमी शब्द को पहले 1949 में पेश किया गया था। ए। सूखे कणिकाओं के अंश के निर्धारण के लिए क्लाउड। बाद में यह पता चला कि माइक्रोसोम का पूरा अंश, जो झिल्ली और कणिकाओं से बना है, प्रोटीन संश्लेषण के लिए जिम्मेदार नहीं है, बल्कि केवल अन्य राइबोन्यूक्लियोप्रोटीन भाग हैं। क्यूई चस्तकी 1958 आर रॉबर्ट्स राइबोसोम द्वारा बुलाया गया था।
1958-1959 में बैक्टीरियल राइबोसोम का क्लासिक अध्ययन ए. टिसियर और जे। वाटसन द्वारा किया गया था। जीवाणु राइबोसोम पौधों और जीवों की तुलना में थोड़े अधिक पाए गए। जे. लिटलटन (1960), एम. क्लार्क (1964) और ई.एन. स्वेटेलो (1966) ने दिखाया कि जीवित पौधों और माइटोकॉन्ड्रिया के क्लोरोप्लास्ट के राइबोसोम बैक्टीरिया के प्रकार से नीचे होते हैं। ए। टिज़ियर और अन्य (1958) ने दिखाया कि राइबोसोम दो तंत्रिका सबयूनिट्स में अलग हो जाते हैं, जिन्हें एक आरएनए अणु द्वारा प्रतिस्थापित किया जा सकता है। उदाहरण के लिए, 1950 के दशक में, यह महत्वपूर्ण था कि राइबोसोमल आरएनए का त्वचा अणु कई छोटे टुकड़ों से बना होता है। प्रोट ए.एस. स्पिरिन, 1960 में पैदा हुए पहले दिखा रहा है कि उप-कणों में आरएनए एक गैर-स्थायी अणु द्वारा दर्शाया गया है। डी. वालर (1960), स्टार्च जेल में अतिरिक्त वैद्युतकणसंचलन के लिए राइबोसोमल प्रोटीन को विभाजित करते हुए, यह स्थापित करते हुए कि बदबू विषम है। अधिकतर, वालर के डेटा के बारे में संदेह होने पर, यह सोचा गया था कि राइबोसोम प्रोटीन सख्ती से सजातीय होने के लिए जिम्मेदार है, उदाहरण के लिए, टीएमवी प्रोटीन। इस घंटे में, डी। वालर, आर। ट्राउट, पी। ट्रुब और अन्य जैव रसायनविदों के शोध के परिणामस्वरूप, यह स्पष्ट हो गया कि प्रोटीन की संरचना के लिए 50 से अधिक बिल्कुल अलग कण राइबोसोमल कणों के गोदाम में प्रवेश कर सकते हैं। 1963 में ए.एस. स्पिरिन राइबोसोमल सबपार्ट्स खुलने के लिए बहुत आगे हैं और दिखाते हैं कि राइबोसोम एक कॉम्पैक्ट रूप से मुड़े हुए राइबोन्यूक्लियोप्रोटीन स्ट्रैंड हैं, जिन्हें गायन दिमाग में उड़ाया जा सकता है। 1967 - 1968 में पीपी। एम. नोमुरा ने उस प्रोटीन के राइबोसोमल आरएनए से जैविक रूप से सक्रिय सबयूनिट का पूरी तरह से पुनर्निर्माण किया और इसे राइबोसोम में पाया, जिसमें आरएनए प्रोटीन विभिन्न सूक्ष्मजीवों से संबंधित थे।
अब तक, राइबोसोमल आरएनए की भूमिका को समझा नहीं गया है। यह माना जाता है कि एक विशिष्ट विशिष्ट मैट्रिक्स है, जिसके लिए राइबोसोमल भाग को ढालते समय, राइबोसोमल प्रोटीन (ए.एस. स्पिरिन, 1968) की संख्या से त्वचा के ठीक उसी स्थान को जानना आवश्यक है।
ए. रिच (1962) ने आईआरएनए के एक स्ट्रैंड द्वारा एक साथ जुड़े कई राइबोसोम के समुच्चय को दिखाया। क्यूई परिसरों को पॉलीसोम कहा जाता था। पॉलीसोम की अभिव्यक्ति ने रिच एंड वॉटसन (1963) को यह स्वीकार करने की अनुमति दी कि पॉलीपेप्टाइड लैंसेट का संश्लेषण राइबोसोम पर होता है, क्योंकि यह iRNA के लैंसेट से होकर गुजरता है। दुनिया में, कण में iRNA के लैंसेट के साथ राइबोसोम को पार करते हुए, प्रोटीन के पॉलीपेप्टाइड लैंसेट की जानकारी पढ़ी जाती है, और लाइन के साथ नए राइबोसोम iRNA के रीड एंड में आते हैं, जो कंपन होता है। रिच और वॉटसन के आंकड़ों से, यह स्पष्ट था कि क्लिटिनी में पॉलीसोम का महत्व प्रोटीन के बड़े पैमाने पर उत्पादन से संबंधित है, जिसके बाद राइबोसोम द्वारा मैट्रिक्स को डेसीलेकॉम में पढ़ा जाता है।
नतीजतन, 1963 - 1970 पीपी में एम। निरेनबर्ग, एस। ओचोआ, एफ। लिपमैन, जी। कुरानी और अन्य का शोध। में। यह स्पष्ट हो गया है कि अनुवाद प्रक्रिया में कई आईआरएनए, राइबोसोम, एटीपी और एमिनोएसिल-टीआरएनए बड़ी संख्या में विभिन्न कारकों का भाग्य लेते हैं, और अनुवाद प्रक्रिया को मानसिक रूप से तीन चरणों में विभाजित किया जा सकता है - दीक्षा, सामान्य अनुवाद और समाप्ति
अनुवाद की शुरुआत का अर्थ है राइबोसोम-मैट्रिक्स पॉलीन्यूक्लियोटाइड-एमिनोएसिल-टीआरएनए कॉम्प्लेक्स में पहले पेप्टाइड लिंकेज का संश्लेषण। यह दीक्षा गतिविधि किसी भी एमिनोएसिल-टीआरएनए में नहीं पाई जाती है, बल्कि औपचारिक मेथियोनील-टीआरएनए में होती है। त्स्या भाषण पहली बार 1964 में देखा गया था। एफ सेंगर और के मार्कर। एस. ब्रेचर और के. मार्कर (1966) ने दिखाया कि फॉर्मिलमेथियोनिल-टीआरएनए के आरंभिक कार्य को स्पोरिडिटी द्वारा राइबोसोम के पेप्टिडाइल केंद्र में बढ़ावा दिया जाता है। कोब पर अनुवाद के लिए, प्रोटीन कारक दीक्षा की गतिविधियाँ और भी महत्वपूर्ण हैं, जो एस। ओचोआ, एफ। ग्रोट और आसपास के अन्य केंद्रों की प्रयोगशालाओं में देखी गई थीं। राइबोसोम में पहला पेप्टाइड लिंकेज स्थापित होने के बाद, अनुवाद शुरू किया जाता है, ताकि बाद में पॉलीपेप्टाइड के सी-टर्मिनस में अमीनोसिल अतिरिक्त जुड़ जाए। प्रसारण प्रक्रिया के बहुत सारे विवरण के। मुनरो और जे। बिशप (इंग्लैंड), आई। रिचलिक और एफ। शोरम (चेकोस्लोवाकिया), एफ। लिपमैन, एम। ब्रेचर, डब्ल्यू। गिल्बर्ट (यूएसए) और अन्य योगदानकर्ता। 1968 आर. ए.एस. स्पिरिन को मूल परिकल्पना का प्रस्ताव देकर राइबोसोम के कार्य के तंत्र की व्याख्या करने के लिए धन्यवाद। निजीकरण मेहनिज़्मोम, जो अनुवाद के घंटे के दौरान टीआरएनए और आईआरएनए आंदोलन के सभी विस्तार को सुनिश्चित करता है, राइबोसोम उप-कणों का आवधिक रोमिंग और ज़मीकन्या है। प्रसारण का अंत मैट्रिक्स में एन्कोड किया गया है, जिसे टर्मिनेटिंग कोड का बदला लेने के तरीके के रूप में पढ़ा जाता है। जैसा कि एस ब्रेनर (1965 - 1967) द्वारा दिखाया गया है, ऐसे कोडन ट्रिपल यूएए, यूएजी और यूजीए हैं। एम. कापेची (1967) ने विशेष प्रोटीन समाप्ति कारकों का भी खुलासा किया। प्रोटीन कारकों की भागीदारी के बिना राइबोसोम (1972 - 1975) में प्रोटीन के तथाकथित "गैर-एंजाइमी" संश्लेषण के विवरण के ए। एस। स्पिरिनिम और एल। पी। गैवरिलोवा। प्रोटीन जैवसंश्लेषण के विकास को समझने के लिए यह बिंदु महत्वपूर्ण है।
जीन और प्रोटीन की गतिविधि का विनियमन
आणविक जीव विज्ञान में पहले क्षेत्र में प्रोटीन संश्लेषण की विशिष्टता की समस्या के बाद, प्रोटीन के संश्लेषण के नियमन की समस्या, मुख्य रूप से, जीन गतिविधि का नियमन दिखाई दिया।
कोशिकाओं के कार्यात्मक गैर-अनुपालन और इसके साथ जुड़े दमन और जीन की सक्रियता लंबे समय से आनुवंशिकीविदों के सम्मान से कमजोर है, लेकिन जीन गतिविधि को नियंत्रित करने के लिए वास्तविक तंत्र अदृश्य हो गया है।
सबसे पहले, हिस्टोन प्रोटीन के विकास में शामिल जीनों की नियामक गतिविधि की व्याख्या करने का प्रयास करें। XX सदी के कोब 40 के दशक में और अधिक दोस्त स्टैडमैन। उन्होंने इस विचार को उठाया कि हिस्टोन इस दुनिया में मुख्य भूमिका निभा सकते हैं। नडाल की बदबू ने हिस्टोन प्रोटीन की रासायनिक प्रकृति के बारे में डेटा की पहली रीडिंग को छीन लिया। निने kіlkіst faktіv, yakі svіdchat savagery tsієї परिकल्पना, त्वचा रॉक daedalі अधिक zrostaє के साथ।
* (ई। स्टेडमैन, ई। स्टेडमैन। कोशिका नाभिक के मूल प्रोटीन।- दार्शनिक। ट्रांस। रॉय। सामाजिक लंदन, 1951, वी. 235, 565 - 595।)
उसी समय, अधिक डेटा जमा हो जाता है, जैसे कि यह कहना कि जीन गतिविधि का नियमन एक बड़े पैमाने पर ढहने वाली प्रक्रिया है, कम सरल है हिस्टोन प्रोटीन के अणुओं के साथ जीन की बातचीत। 1960 - 1962 में आरआर। आर बी खेसीन-लूर की प्रयोगशाला में, यह निर्धारित किया गया था कि फेज के जीन को अलग-अलग समय पर गिना जा सकता है: टी 2 फेज के जीन को प्रारंभिक चरण में जोड़ा जा सकता है, जिसका कामकाज पहली पंक्ति में किया गया था। जीवाणु कोशिकाओं के संक्रमण, और आरएनए को प्रारंभिक जीन के पूरा होने के बाद संश्लेषित किया गया था।
1961 में फ्रांसीसी जैव रसायनज्ञ एफ। जैकब और जे। मोनोड ने जीन गतिविधि के नियमन की योजना का प्रचार किया, जिसने कोशिकाओं के नियामक तंत्र के विकास में महत्वपूर्ण भूमिका निभाई। जैकब और मोनोड की योजना के अनुसार, डीएनए में संरचनात्मक (सूचनात्मक) जीन भी जीन-नियामक और जीन-ऑपरेटर होते हैं। नियामक जीन विशिष्ट भाषण के संश्लेषण को एन्कोड करता है - एक दमनकारी, जिसे प्रारंभ करनेवाला और ऑपरेटर जीन दोनों से जोड़ा जा सकता है। संरचनात्मक जीन के साथ जंजीरों के जीन-ऑपरेटर, और जीन-रेगुलेटर को एक ही रिमोट वाले में स्थानांतरित किया जाता है। मध्य प्रारंभ करनेवाला के रूप में, उदाहरण के लिए, लैक्टोज, फिर जीन-नियामक द्वारा संश्लेषित, दमनकर्ता जीन-ऑपरेटर i से बांधता है, इसे अवरुद्ध करता है, पूरे ऑपेरॉन के काम की नकल करता है (संचालक के साथ एक बार संरचनात्मक जीन का एक ब्लॉक) उन्हें ले जाता है)। उनके दिमाग में एंजाइम का कोई संकल्प नहीं है। यदि मध्य एक इंड्यूसर (लैक्टोज) है, तो रेगुलेटर जीन का उत्पाद - रेप्रेसर - लैक्टोज के साथ प्रतिक्रिया करेगा और ऑपरेटर जीन को ब्लॉक करेगा। आप किस दिशा में हैं सक्षम रोबोटसंरचनात्मक जीन जो एंजाइम के संश्लेषण को कूटबद्ध करता है, वह एंजाइम (लैक्टोज) माध्यम में घुल जाता है।
जैकब और मोनॉड के अनुसार, विनियमन योजना सभी अनुकूली एंजाइमों के लिए स्थिर है और यदि प्रतिक्रिया उत्पाद की अधिकता से एंजाइम को ध्यान में रखा जाता है, और प्रेरण के दौरान, यदि सब्सट्रेट की शुरूआत एंजाइम के संश्लेषण का कारण बनती है, तो इसे दबाया जा सकता है। . जीन की गतिविधि के और नियमन के लिए 1965 में जैकब और मोनो को सम्मानित किया गया। नोबेल पुरुस्कार।
हाथ के पीछे, यह योजना दूर की कौड़ी को दे दी गई थी। हालांकि, यह दिखाया गया है कि इस सिद्धांत के अनुसार जीन का नियमन केवल बैक्टीरिया में ही नहीं, बल्कि अन्य जीवों में भी संभव है।
1960 में शुरू आणविक जीव विज्ञान में महत्वपूर्ण स्थान जीनोम के निम्नलिखित संगठन और यूकेरियोटिक जीवों में क्रोमैटिन की संरचना द्वारा कब्जा कर लिया गया है (जे। बोनर, आर। ब्रितन, डब्ल्यू। ओल्फ्री, पी। वॉकर, यू। एस। चेन्त्सोव, आई। बी। ज़बर्स्की एट इन। ) प्रतिलेखन विनियमन (ए। मिर्स्की, जी। पी। जॉर्जीव, एम। बर्नस्टील, डी। गोल, आर। त्सनेव, आर। आई। सालगानिक)। लंबे समय तक दमन करने वाले का अपरिचित स्वभाव पीछे छूट गया। 1968 आर. Ptashne (यूएसए) दिखा रहा है कि प्रोटीन एक दमनकारी है। वेन ने जे. वाटसन की प्रयोगशाला में योगो को देखा और पाया कि दमनकारी, जाहिरा तौर पर, प्रारंभ करनेवाला (लैक्टोज) के लिए sporidnіst हो सकता है और तुरंत लाख-ओपेरॉन के जीन-ऑपरेटर को "जान" सकता है और विशेष रूप से इसके साथ जुड़ सकता है।
शेष 5-7वें वर्षों में, जीन गतिविधि के एक और महत्वपूर्ण मध्य - प्रवर्तक की उपस्थिति के बारे में डेटा हटा दिया गया था। यह पता चला कि, ऑपरेटर के बोर्ड की प्रकृति के अनुसार, उत्पाद आने से पहले, जीन-नियामक पर संश्लेषण करना - दमनकर्ता का प्रोटीन भाषण, एक और साजिश है, साथ ही साथ नियामक प्रणाली के सदस्यों के लिए एक लिंक भी है। जीन गतिविधि। एंजाइम आरएनए पोलीमरेज़ के लिए एक प्रोटीन अणु पौधे के अंत में आता है। प्रमोटर डिवीजन डीएनए में न्यूक्लियोटाइड के अद्वितीय अनुक्रम और आरएनए पोलीमरेज़ प्रोटीन के विशिष्ट विन्यास को पारस्परिक रूप से पहचान सकता है। आनुवंशिक जानकारी को पढ़ने की अंतर्निहित प्रक्रिया की मान्यता की दक्षता के कारण जीन ऑपेरॉन का अनुक्रम दिया गया है, जो प्रमोटर से जुड़ा हुआ है।
क्रीम का वर्णन जैकब और मोनो योजनाओं द्वारा किया गया है, क्लिनी में, जीन के नियमन के अन्य तंत्र पाए जाते हैं। एफ. जैकब और एस. ब्रेनर (1963) ने स्थापित किया कि जीवाणु डीएनए की प्रतिकृति के नियमन को कोशिकीय झिल्ली द्वारा एकल रैंक द्वारा नियंत्रित किया जाता है। जैकब (1954) के अध्ययन ने विभिन्न प्रचारों को शामिल करने पर बार-बार दिखाया है कि, लाइसोजेनिक बैक्टीरिया की कोशिकाओं में विभिन्न उत्परिवर्तजन कारकों के प्रवाह के तहत, प्रोफ़ेज जीन की प्रतिकृति का चयन शुरू किया जाता है, और जीनोम की प्रतिकृति होती है। मास्टर अवरुद्ध है। 1970 आर. एफ. बेल ने दिखाया कि छोटे डीएनए अणु कोशिका द्रव्य से नाभिक तक जा सकते हैं और वहां स्थानांतरित हो सकते हैं।
इस प्रकार, जीन गतिविधि का नियमन समान प्रतिकृति, प्रतिलेखन और अनुवाद से प्रभावित हो सकता है।
एंजाइमों के संश्लेषण और उनकी गतिविधि दोनों के नियमन में महत्वपूर्ण सफलताएँ प्राप्त हुई हैं। क्लिटिन में एंजाइम गतिविधि के नियमन की घटना पर, ए। नोविक और एल। स्ज़ीलार्ड ने 50 के दशक में दिखाया। जी. उम्बर्गर (1956) ने स्थापित किया कि क्लिटिनी में चलटल लिगामेंट के लिए प्रतिक्रियाओं के लांसर के टर्मिनल उत्पाद के साथ एंजाइम की गतिविधि को दबाने का एक और भी अधिक तर्कसंगत तरीका है। जैसा कि जे। मोनोड, जे। चेंज, एफ। जैकब, ए। परदित और अन्य (1956 - 1960) द्वारा स्थापित किया गया था, एंजाइम गतिविधि का विनियमन एलोस्टेरिक सिद्धांत का पालन कर सकता है। एंजाइम या तो इसके उप-इकाइयों में से एक है, सब्सट्रेट के लिए इसकी स्पोरिडिटी के अलावा, यह प्रतिक्रिया उत्पादों में से एक के साथ स्पोरिडेनिस्ट हो सकता है। इस तरह के उत्पाद-संकेत के प्रवाह के तहत, एंजाइम अपनी रचना को बदल देता है, जो गतिविधि को प्रेरित करता है। नतीजतन, एंजाइमी प्रतिक्रियाओं का पूरा भाला सिल पर ही अंकित हो जाता है। डी. विमेंटा आर. वुडवर्ड (1952; नोबेल पुरस्कार विजेता, 1965) ने एंजाइमी प्रतिक्रियाओं में और स्पष्ट अर्थों में और एक एलोस्टेरिक प्रभाव की उपस्थिति में प्रोटीन के गठनात्मक परिवर्तनों की भूमिका की ओर इशारा किया।
प्रोटीन की संरचना और कार्य
नतीजतन, टी। ओसबोर्न, जी। हॉफमेस्टर, ए। गुरबर, एफ। शुल्ज और अन्य का काम उदाहरण के लिए XIXमें। एक क्रिस्टल लुक में अमीर जीव और रोजलिन गिलहरी को ले जाया गया। लगभग उसी समय, विभिन्न भौतिक विधियों की सहायता के लिए, कुछ प्रोटीनों के आणविक प्रोटीन स्थापित किए गए थे। तो, 1891 में। ए. सबनीव और एन. अलेक्जेंड्रोव ने बताया कि ओवलब्यूमिन का आणविक भार 14,000 था; 1905 में ई. रीड ने यह स्थापित किया कि हीमोग्लोबिन का आणविक भार 48,000 है। प्रोटीन की बहुलक संरचना की खोज 1871 में हुई थी। जी. ग्लेज़िवेट्स और डी. गैबरमैन। प्रोटीन में अमीनो एसिड अवशेषों के पेप्टाइड लिंकेज के बारे में विचार टी। कर्टियस (1883)। अमीनो एसिड के रासायनिक संघनन पर काम करता है (ई। शाल, 1871; जी। शिफ, 1897; एल। बालबियानो और डी। ट्रैसियाट्टी, 1900) और हेटरोपॉलीपेप्टाइड्स के संश्लेषण (ई। फिशर, 1902 - 1907, प्रोटीन की रासायनिक संरचना।
पहला क्रिस्टल एंजाइम (यूरेस) 1926 में हटा लिया गया था। जे. सुमनेर (नोबेल पुरस्कार, 1946), और 1930 पी। जे. नॉर्थ्रॉप (नोबेल पुरस्कार, 1946) ने क्रिस्टल पेप्सिन लिया। मेरे शोध के बाद, यह स्पष्ट हो गया कि एंजाइम प्रोटीन की प्रकृति को बदल सकते हैं। 1940 में एम. कुनित्स ने क्रिस्टल RNase देखा। 1958 तक, पहले से ही 100 से अधिक क्रिस्टलीय एंजाइम और 500 से अधिक एंजाइम थे जिन्हें गैर-क्रिस्टलीय रूप में देखा गया था। व्यक्तिगत प्रोटीन में उच्च शुद्धता की तैयारी की सामग्री ने उनकी प्राथमिक संरचना और मैक्रोमोलेक्यूलर संगठन की व्याख्या की।
मनुष्यों के आणविक जीव विज्ञान और आनुवंशिकी के विकास का बहुत महत्व है, विशेष रूप से एल। पॉलिंग (1940) ने एक असामान्य हीमोग्लोबिन एस देखा, जो गंभीर बीमारी वाले लोगों के एरिथ्रोसाइट्स में देखा जाता है - सिकल सेल एनीमिया। 1955 - 1957 में आरआर। वी। इंग्रेम विकोरिस्टोवुव एफ। सेंगर की "उंगलियों की उंगलियों" की विधि (आग की लपटें, कागज पर क्रोमैटोग्राफी के दौरान ओकेमी पेप्टाइड्स द्वारा अनुमोदित) घास के मैदान और ट्रिप्सिन के साथ हीमोग्लोबिन एस के हाइड्रोलिसिस के उत्पादों के विश्लेषण के लिए। 1961 में इनग्राम ने दिखाया कि हीमोग्लोबिन एस सामान्य हीमोग्लोबिन में केवल एक अमीनो एसिड की अधिकता की प्रकृति के कारण पाया जाता है: सामान्य हीमोग्लोबिन में लांस स्थिति में ग्लूटामिक एसिड की अधिकता होती है, और हीमोग्लोबिन एस में वेलिन की अधिकता होती है। पॉलिंग के इस स्वीकारोक्ति से फिर से (1949) इस बात की पुष्टि हुई कि सिकल-क्लिटिन एनीमिया एक आणविक प्रकृति की बीमारी है। स्पैडकोव के हीमोग्लोबिन मैक्रोमोलेक्यूल के आधे हिस्से में त्वचा में अमीनो एसिड की सिर्फ एक अतिरिक्त परिवर्तन इस तथ्य को जन्म दे सकता है कि हीमोग्लोबिन खट्टा की कम सांद्रता पर आसानी से संक्रमित हो जाता है और क्रिस्टलीकरण करना शुरू कर देता है, जिससे कोशिकाओं की संरचना को नुकसान होता है। इन अध्ययनों ने स्पष्ट रूप से दिखाया है कि प्रोटीन की संरचना एक एकल अमीनो एसिड अनुक्रम है, जैसा कि जीनोम में एन्कोड किया गया है। K. Anfinsen (1951) के लेखकों ने मैक्रोमोलेक्यूल की एक अद्वितीय जैविक रूप से सक्रिय संरचना के निर्माण में प्रोटीन की प्राथमिक संरचना के महत्व को नोट किया। Anfіnsen scho bіologіchno सक्रिय मैक्रोस्ट्रक्चर pankreatichnoї ribonuclease scho vtrachaєtsya दिखा रहा है rezultatі vіdnovlennya, zumovlena amіnokislotnoyu poslіdovnіstyu i Mauger znovu vinikati सहज रूप से okislennі के znovu vinikati।
वर्तमान समय तक, बड़ी संख्या में एंजाइमों के तंत्र और समृद्ध प्रोटीन की संरचना का विस्तार से निर्धारण किया गया है।
1953 आर. एफ। सेंगर ने इंसुलिन के अमीनो एसिड अनुक्रम की स्थापना की। : यह प्रोटीन दो पॉलीपेप्टाइड लैंसेट से बना होता है, जो दो डाइसल्फ़ाइड क्रॉसलिंक्स से जुड़ता है। एक लांसर के पास कुल 21 अमीनो एसिड सरप्लस हैं, और दूसरे में - 30 सरप्लस हैं। 10 roki के बारे में budіvlі tsgogo pіvnya सरल प्रोटीन सेंगर विट्रेटिव की व्याख्या पर। 1958 पी. इस काम के लिए उन्हें नोबेल पुरस्कार से सम्मानित किया गया था। डब्ल्यू। स्टीन और एस। मूर (1957) द्वारा एक स्वचालित अमीनो एसिड विश्लेषक के निर्माण के बाद, प्रोटीन के आंशिक हाइड्रोलिसिस के उत्पादों की पहचान में काफी तेजी आई है। 1960 आर. स्टीन और मूर ने उसे पहले ही बता दिया था। वह 124 अमीनो एसिड अवशेषों के किसी भी प्रतिनिधित्व के पेप्टाइड लांस, राइबोन्यूक्लीज के अनुक्रम को कैसे नामित कर सकता है। इसके अलावा, टुबिंगन (एफआरएन) एफ। एंडरर और अन्य में जी। श्राम की प्रयोगशाला में, उन्होंने टीएमवी प्रोटीन में एमिनो एसिड अनुक्रम निर्धारित किया। तब अमीनो एसिड अनुक्रम की पहचान मानव मायोग्लोबिन (ए। एडमन्सन) और α- और मानव हीमोग्लोबिन में β-लांस (जी। ब्रूनिट्जर, ई। श्रोएडर एट अल।), चिकन अंडे के सफेद से लाइसोजाइम (जे। जोलेट, डी। कीफील्ड)। 1963 पी. एफ। शोरम और बी। कील (चेकोस्लोवाकिया) ने केमोट्रिप्सिनोजेन अणु में अमीनो एसिड के अनुक्रम की स्थापना की। उसी रोसी में, ट्रिप्सिनोजेन के अमीनो एसिड अनुक्रम को सौंपा गया था (एफ। शोर, डी। वॉल्श)। 1965 में के. ताकाहाशी ने टी1 राइबोन्यूक्लीज की प्राथमिक संरचना की स्थापना की। तब अमीनो एसिड का क्रम प्रोटीन की संख्या में अधिक निर्धारित किया गया था।
जैसा कि यह पता चला है, tієї chi inshої संरचना संश्लेषण के पदनाम की शुद्धता का अवशिष्ट प्रमाण। 1969 में आर मेरिफिल्ड (यूएसए) ने अग्नाशयी राइबोन्यूक्लिअस के रासायनिक संश्लेषण का बीड़ा उठाया। संश्लेषण विधि के अलावा उन्होंने एक ठोस-चरण मेरिफिल्ड वाहक पर विकसित किया, एक के बाद एक अमीनो एसिड को लैंसेट में जोड़ते हुए, उसी क्रम तक, जैसा कि स्टीन और मूर द्वारा वर्णित है। नतीजतन, मैंने प्रोटीन को हटा दिया, जो इसके गुणों में अग्नाशयी राइबोन्यूक्लिएज ए के समान था। वी। स्टीन, एस। मूर और के। एनफिन्सन को राइबोन्यूक्लिज की खोज के लिए, तुला का जन्म 1972 में हुआ था। नोबेल पुरस्कार से सम्मानित किया गया। प्राकृतिक प्रोटीन का यह संश्लेषण भविष्य में नियोजित अनुक्रम तक किसी भी प्रोटीन के निर्माण की संभावना की ओर इशारा करते हुए भव्य संभावनाओं को दर्शाता है।
एक्स-रे विवर्तन अध्ययनों से, डब्ल्यू। एस्टबरी (1933) ने दिखाया कि प्रोटीन अणुओं के पेप्टाइड लेंस एक सख्त गायन-समान क्रम में मुड़ या व्यवस्थित होते हैं। इस समय से, बहुत सारे लेखक सफेद डोरी बिछाने के तरीकों के बारे में अलग-अलग परिकल्पनाओं के साथ आए, और 1951 तक, सभी मॉडल सट्टा उद्देश्यों से भरे हुए थे, जो प्रयोगात्मक डेटा से मेल नहीं खाते थे। 1951 आर. एल। पॉलिंग और आर। कोरी ने शानदार कार्यों की एक श्रृंखला प्रकाशित की, जिसमें प्रोटीन की माध्यमिक संरचना का सिद्धांत तैयार किया गया था - α-हेलिक्स का सिद्धांत। एक पंक्ति में, यह स्पष्ट हो गया कि प्रोटीन एक अधिक तृतीयक संरचना बनाते हैं: पेप्टाइड लैंसेट के α-हेलिक्स को एक रैखिक क्रम में मोड़ा जा सकता है, जिससे एक कॉम्पैक्ट संरचना प्राप्त करना संभव हो जाता है।
1957 आर. जे. केंड्रू और योग विशेषज्ञों ने सबसे पहले मायोग्लोबिन संरचना के त्रिविमर मॉडल का प्रचार किया। इस मॉडल को तब दशकों तक खींचकर परिष्कृत किया गया था, जब तक कि 1961 में प्रोटीन की अंतरिक्ष संरचना की विशेषता के साथ एक सबबैग रोबोट दिखाई नहीं दिया। 1959 आर. एम. पेरुट्ज़ और स्पाइवरोबिट्निकी ने हीमोग्लोबिन की त्रिविमिर संरचना की स्थापना की। इस काम के लिए, श्रमिकों ने 20 से अधिक वर्षों का समय बिताया (हीमोग्लोबिन की पहली एक्स-रे 1937 में पैदा हुए पेरुट्ज़ द्वारा ली गई थी)। चूंकि हीमोग्लोबिन अणु कई उप-इकाइयों से बना है, इसलिए, इस संगठन को समझने के बाद, पेरुट्ज़ सिम ने पहले प्रोटीन की चौथाई संरचना का वर्णन किया। 1962 में केंड्रू और पेरुट्ज़ में प्रोटीन की तुच्छ संरचना को डिजाइन करने के कार्य के लिए। नोबेल पुरस्कार से सम्मानित किया गया था।
पेरू ने हीमोग्लोबिन की संरचना के एक अंतरिक्ष मॉडल के निर्माण की अनुमति दी। इस प्रोटीन के कामकाज के तंत्र की समझ के करीब पहुंचें, जो कि ऐसा लगता है, भगशेफ जीवों में खटास के हस्तांतरण का कारण बनता है। योजना 1937 आर. एफ। गौरोविट्ज ने इस तथ्य के साथ एक संबंध विकसित किया कि खट्टे के साथ हीमोग्लोबिन की बातचीत फिर से प्रोटीन की संरचना में बदलाव के साथ हो सकती है। 1960 के दशक में, पेरुट्ज़ और योग शोधकर्ताओं ने ऑक्सीजन ऑक्सीकरण के बाद हीमोग्लोबिन में एक मामूली बदलाव का खुलासा किया, जिसने एसिड के साथ लिंक के बाद हवा में परमाणुओं के विनाश का आह्वान किया। इस आधार पर, प्रोटीन मैक्रोमोलेक्यूल्स के "श्वास" के बारे में बयान तैयार किए गए थे।
1960 आर. D. फिलिप्स और योगो स्पाइवोलॉजिस्ट ने लाइसोजाइम अणु के एक्स-रे विवर्तन विश्लेषण की पहचान की है। 1967 तक वह कमोबेश इस प्रोटीन के संगठन और एक अणु में चार परमाणुओं के स्थानीयकरण का विवरण स्थापित करने में सक्षम था। Krym ts'ogo, Philips z'yasuvav सब्सट्रेट (triacetylglucosamine) के लिए लाइसोजाइम के आगमन की प्रकृति। Tse ने इस एंजाइम के तंत्र को बहाल करने की अनुमति दी। इस प्रकार, उस मैक्रोमोलेक्यूलर संगठन की प्राथमिक संरचना के ज्ञान ने न केवल विभिन्न एंजाइमों के सक्रिय केंद्रों की प्रकृति को स्थापित करना संभव बना दिया, बल्कि इन मैक्रोमोलेक्यूल्स के कामकाज के तंत्र को भी प्रकट करना संभव बना दिया।
इलेक्ट्रॉन माइक्रोस्कोपी में विधियों की पसंद ने ऐसे तह प्रोटीन समाधानों के मैक्रोमोलेक्यूलर संगठन के सिद्धांतों को प्रकट करने में मदद की, जैसे धागे से कोलेजन, फाइब्रिनोजेन, m'yazyv के तेजी से बढ़ने वाले तंतु और इन। उदाहरण के लिए, 1950 के दशक में, तेजी से चलने वाले उपकरण का एक मॉडल प्रस्तावित किया गया था। विनयटकोव के लिए, म्यूकोसल डिसरिथिमिया के तंत्र को समझने के महत्व को यू.ए. एंगेलगार्ड और एम.एम. हुसिमोवा (1939) मायोसिन की एटीपी-एज़ गतिविधि द्वारा बहुत कम मान्यता प्राप्त है। Tse का मतलब था कि m'yazovy स्थिरता के कार्य का आधार भौतिक और रासायनिक शक्तियों के परिवर्तन और एडेनोसिन ट्राइफॉस्फोरिक एसिड (डिव। धारा 11) के प्रवाह के तहत अल्पकालिक प्रोटीन के मैक्रोमोलेक्यूलर संगठन में निहित है।
तह जैविक संरचनाओं के सिद्धांतों को समझने के लिए, छोटे वायरोलॉजिकल अध्ययनों का महत्व महत्वपूर्ण है (div. Rozdil 25)।
अदृश्य समस्याएं
आधुनिक आणविक जीव विज्ञान में मुख्य सफलताएं न्यूक्लिक एसिड ग्राफ्टिंग के परिणामों में अधिक महत्वपूर्ण रूप से प्राप्त की गई हैं। virіshenі की सभी समस्याओं से दूर tsіy galuzі sche पर प्रोट नेविट। ग्रेट ज़ुसिल विमागाटाइम, ज़ोक्रेमा, जीनोम के पूरे न्यूक्लियोटाइड अनुक्रम को समझने वाला। यह समस्या डीएनए विषमता की समस्या से अटूट रूप से जुड़ी हुई है और क्लिटिन की कुल आनुवंशिक सामग्री से अलग-अलग अणुओं के विभाजन और पहचान के नए गहन तरीकों के विकास के कारण है।
सिच पीर ज़ुसिल से पहले, यह मुख्य रूप से प्रोटीन और न्यूक्लिक एसिड के ओकेरेमू किण्वन पर केंद्रित था। कोशिकाओं में, बायोपॉलिमर असंगत रूप से एक से एक से जुड़े होते हैं और न्यूक्लियोप्रोटीन के रूप में शीर्ष रैंक के रूप में कार्य करते हैं। उसी समय, एक विशेष गंभीरता के साथ, प्रोटीन और न्यूक्लिक एसिड के संयोजन की आवश्यकता का पता चला था। सबसे आगे प्रोटीन द्वारा न्यूक्लिक एसिड की पहचान की समस्या है। पहले से ही, क्रोमोसोम, राइबोसोम और अन्य संरचनाओं की किसी भी स्पष्ट संरचना और कार्यों के बिना, बायोपॉलिमर की इस तरह की अन्योन्याश्रयता के विकास के लिए कदम उठाए गए हैं। इसके बिना, जीन गतिविधि के नियमन को समझना और रोबोटिक संश्लेषण तंत्र के सिद्धांतों को समझना भी असंभव है। जैकब और मोनोड के काम के बाद, परमाणु सामग्री के संश्लेषण में झिल्ली के नियामक महत्व के बारे में नया डेटा सामने आया। उद्देश्य डीएनए प्रतिकृति के नियमन में झिल्ली की भूमिका की सबसे बड़ी जांच का कार्य निर्धारित करना है। सामान्य तौर पर, जीन गतिविधि और सेलुलर गतिविधि के नियमन की समस्या आधुनिक आणविक जीव विज्ञान की सबसे महत्वपूर्ण समस्याओं में से एक बन गई है।
आधुनिक बायोफिजिक्स कैंप
आणविक जीव विज्ञान की समस्याओं के साथ लिंक पर बायोफिज़िक्स का विकास है। जीव विज्ञान के क्षेत्र में रुचि उत्तेजक है, एक तरफ, शरीर में विभिन्न प्रकार के क्षरणों के सार्वभौमिक टीकाकरण की आवश्यकता है, और दूसरी ओर, जीवन के अतिरिक्त भौतिक और भौतिक और रासायनिक आधारों, आणविक कारकों की आवश्यकता है।
नई सूक्ष्म भौतिक और रासायनिक विधियों के विकास के परिणामस्वरूप आणविक संरचना और उनमें शामिल प्रक्रियाओं के बारे में सटीक जानकारी प्राप्त करना संभव हो गया है। इलेक्ट्रोकैमिस्ट्री की पहुंच के आधार पर, आयन-चयनात्मक इलेक्ट्रोड (जी। ईज़ेनमैन, बी.पी. निकोल्स्की, खुरी, 50 - 60 वर्ष) का उपयोग करके जैव-विद्युत क्षमता के विमिरुवन्न्या की विधि को पूरी तरह से विकसित किया गया था। इंफ्रारेड स्पेक्ट्रोस्कोपी (विभिन्न लेजर उपकरणों के साथ) के अभ्यास में डेडल्स का अधिक व्यापक रूप से उपयोग किया जाता है, जो प्रोटीन के गठनात्मक परिवर्तन (आई। प्लॉटनिकोव, 1940) को जारी रखने की अनुमति देता है। इसके अलावा, इलेक्ट्रॉन पैरामैग्नेटिक रेजोनेंस की विधि (ई.के. ज़ावोस्की, 1944) और बायोकेमोल्यूमिनसेंट विधि (बी.एन. तारुसोव एट अल।, 1960) ऑक्साइड प्रक्रियाओं के दौरान इलेक्ट्रॉनों के परिवहन का न्याय करने के लिए, ज़ोक्रेमा को अनुमति देती है।
1950 के दशक तक, बायोफिज़िक्स ने एक सैन्य शिविर जीता। योग्य विशेषज्ञों के प्रशिक्षण की आवश्यकता को दोष दें। 1911 के निकट यक्षो यूरोप में, पेक विश्वविद्यालय में, उगोर क्षेत्र में, बायोफिज़िक्स का एक विभाग था, फिर 1973 तक। ऐसे विभाग सभी महान विश्वविद्यालयों में स्थापित हैं।
1960 आर. इंटरनेशनल एसोसिएशन ऑफ बायोफिजिसिस्ट द्वारा आयोजित किया गया था। 1961 में पैदा हुए तोरिशनी सिकल पहली अंतर्राष्ट्रीय बायोफिजिकल कांग्रेस स्टॉकहोम के पास आयोजित की गई थी। 1965 में एक और कांग्रेस हुई। पेरिस में, तीसरा - 1969 में। बोस्टन में, क्वार्टर - 1972 में। मास्को में।
बायोफिज़िक्स में, स्पष्ट कट के लिए दो अलग-अलग लोगों के बीच स्पष्ट अलगाव होता है - आणविक बायोफिज़िक्स और क्लिटिन बायोफिज़िक्स। त्से सीमांकन otrimu y organizatsiyny viraz: इन दो सीधे बायोफिज़िक्स के okremі विभाग बनाए जा रहे हैं। मॉस्को विश्वविद्यालय में, 1953 में बायोफिज़िक्स का पहला विभाग बनाया गया था। जीव विज्ञान और मृदा विज्ञान के संकाय में, और भौतिकी के संकाय में बायोफिज़िक्स विभाग में। उसी सिद्धांत का पालन करते हुए, अमीर और अन्य विश्वविद्यालयों में विभागों का आयोजन किया गया।
आणविक बायोफिज़िक्स
दुनिया के बाकी हिस्सों में, आणविक बायोफिज़िक्स और आणविक जीव विज्ञान के बीच की कड़ी अधिक से अधिक महत्वपूर्ण होती जा रही है, और साथ ही उनके बीच अंतर करना भी महत्वपूर्ण है। मंदी की जानकारी की समस्या पर एक सामान्य हमले में, बायोफिज़िक्स और आणविक जीव विज्ञान के बीच ऐसा सहयोग अपरिहार्य है।
न्यूक्लिक एसिड भौतिकी के अंतिम रोबोटिक विकास में सीधे शीर्षक डीएनए और आरएनए है। अधिक विधियों के महत्व के विकास और एक्स-रे विवर्तन विश्लेषण के उपयोग ने न्यूक्लिक एसिड की आणविक संरचना की व्याख्या की। इस समय गुलाब में इन अम्लों के व्यवहार के विकास पर गहन शोध किया जा रहा है। विशेष रूप से सम्मान किसी भी गठनात्मक संक्रमण "सर्पिल-कॉइल" से जुड़ा होता है, जो चिपचिपाहट, ऑप्टिकल और विद्युत संकेतों में परिवर्तन के साथ होता है। उत्परिवर्तन के तंत्र के विकास के संबंध में, पौधों में न्यूक्लिक एसिड के व्यवहार के साथ-साथ वायरस और फेज के न्यूक्लिक एसिड पर आयनकारी विकिरण का और विकास विकसित होता है। सार्वभौमिक विश्लेषण को पराबैंगनी कंपन की आमद द्वारा समर्थित किया गया था, इतनी अच्छी मिट्टी के वर्णक्रमीय कूड़े के बधिर न्यूक्लिक एसिड हैं। इस तरह के अध्ययनों में इलेक्ट्रॉन पैरामैग्नेटिक रेजोनेंस की विधि द्वारा न्यूक्लिक एसिड और प्रोटीन के सक्रिय रेडिकल्स का पता लगाना बहुत महत्वपूर्ण है। Zastosuvannyam tsgogo vyyazano से सीधे एक पूरे स्वतंत्र को दोष देना।
प्रोटीन संश्लेषण के दौरान डीएनए और आरएनए सूचना कोडिंग और ट्रांसमिशन की समस्या लंबे समय से आणविक बायोफिज़िक्स में एक समस्या रही है, और भौतिकविदों ने इस ड्राइव और अन्य माइक्रोस्कोपी (ई। श्रोडिंगर, जी। गामो) के कारण बार-बार काम किया है। आनुवंशिक कोड की व्याख्या करने के लिए डीएनए हेलिक्स की संरचना पर संख्यात्मक सैद्धांतिक और प्रायोगिक अध्ययन, फोर्जिंग और ट्विस्टिंग थ्रेड्स की व्यवस्था, इन प्रक्रियाओं में भाग लेने वाले भौतिक बलों की घुमाव की आवश्यकता होती है।
अतिरिक्त एक्स-रे विवर्तन विश्लेषण के साथ प्रोटीन अणुओं की संरचना के विकास में आणविक जीव विज्ञान पर महत्वपूर्ण रूप से आणविक बायोफिज़िक्स की सहायता करना, जो पहले 1930 के दशक में स्थापित किया गया था। जे बर्नाल। जैव रासायनिक (एंजाइमी विधियों) के साथ संयुक्त रूप से अलग-अलग भौतिक तरीकों के परिणामस्वरूप, कई प्रोटीनों में आणविक संरचना और अमीनो एसिड फैलाव के अनुक्रम का पता चला था।
वर्तमान इलेक्ट्रॉन-सूक्ष्म अध्ययन, जिसने फोल्डिंग मेम्ब्रेन सिस्टम के क्लिटिन और ऑर्गेनेल में उपस्थिति का खुलासा किया, ने आणविक दिमाग की खोज को प्रेरित किया (डिव। अध्याय 10 और 11)। झिल्लियों के रासायनिक गोदाम का अस्तित्व, जो कि, ज़ोक्रेमा, उनके लिपिड की शक्ति विकसित की जा रही है। Bulo z'yazovan, scho लैंसेग ऑक्सीकरण (यू। ए। वोलोडिमिरोव और एफ। एफ। लिट्विन, 1959; बी। एन। टारसोव और इन।, 1960; आई। आई। इवानोव, 1967) के ओवरऑक्सीडेशन और गैर-एंजाइमी प्रतिक्रियाओं के लिए zdatnі रहते हैं, जो विनाश की ओर ले जाते हैं। झिल्ली कार्य। झिल्लियों के निर्माण के लिए, गणितीय मॉडलिंग (वी। टीएस। प्रेसमैन, 1964 - 1968; एम। एम। शेम्याकिन, 1967; यू। ए। ओविचिनिकोव, 1972) के तरीकों का उपयोग करके झिल्ली की संरचना को भी ठीक किया जाने लगा।
क्लिटिना बायोफिज़िक्स
बायोफिज़िक्स के इतिहास में एक महत्वपूर्ण विकास 1950 के दशक में जैविक प्रक्रियाओं के थर्मोडायनामिक्स के बारे में स्पष्ट बयानों का गठन था, जिसके बाद थर्मोडायनामिक्स के एक और कानून के बावजूद जीवित कोशिकाओं में आत्मनिर्भर ऊर्जा की संभावना के लिए एक अवशिष्ट भत्ता था। जैविक प्रणालियों में Rozuminnya dії tsgogo कानून pov'yazane isz zaprovadzhennyam belgіyskimi ucheniyi । प्रिगोगिन (1945) प्रिगोगिन ने दिखाया कि थर्मोडायनामिक्स के एक अन्य नियम के अनुसार कार्य प्रक्रियाओं के दौरान जीवित कोशिकाओं में सकारात्मक एन्ट्रापी स्थापित होती है। उनके द्वारा प्रस्तुत, समान को मन द्वारा नियुक्त किया गया था, कुछ मदिरा के लिए, तथाकथित स्थिर शिविर (पहले, इसे गतिशील समान भी कहा जाता था), जिसके लिए बहुत सारी मुक्त ऊर्जा (नेगेंट्रॉपी) होती है, जो होनी चाहिए क्लिटिन z zhey, vitraten . के लिए क्षतिपूर्ति इसका कारण क्लिटिन के बाहरी और भीतरी मध्य के बीच अपरिचित संबंध के बारे में एक मौलिक जैविक विचार था। वोनो ने मॉडलिंग पद्धति (ए। बर्टन, 1939; ए। जी। पासिन्स्की, 1967) सहित जीवित प्रणालियों के ऊष्मप्रवैगिकी के वास्तविक विकास की नींव रखी।
* (महत्वपूर्ण प्रणालियों के वैश्विक सिद्धांत को पहली बार 1932 में एल. बर्टलान्फी द्वारा प्रदर्शित किया गया था।)
बायोथर्मोडायनामिक्स के मूल सिद्धांत के आधार पर, जैव रासायनिक प्रक्रियाओं के विकास में जीवन के आवश्यक बौद्धिक आधार को स्थिर दिखाया गया है, जिसके विकास के लिए भाषण के आदान-प्रदान की संख्यात्मक प्रतिक्रियाओं का समन्वय आवश्यक है। नए बायोफिजिकल थर्मोडायनामिक्स के आधार पर, विनिक सीधे, जो समान और आंतरिक कारकों को देखता है, याक यह सुनिश्चित करता है कि प्रतिक्रियाओं का समन्वय स्थिर है। शेष दो दशकों के लिए, अवरोधकों और विशेष रूप से एंटीऑक्सिडेंट (बी। एन। तारुसोव और ए। आई। ज़ुरावलेव, 1954, 1958) की प्रणाली की स्थिर स्थिति के विकास में एक महान भूमिका दिखाई गई। यह स्थापित किया गया है कि स्थिर विकास की श्रेष्ठता बाहरी वातावरण (तापमान) के कारकों और कोशिकाओं के पर्यावरण की भौतिक और रासायनिक शक्ति से जुड़ी है।
बायोथर्मोडायनामिक्स के वर्तमान सिद्धांतों ने हमें अनुकूलन के तंत्र को एक भौतिक और रासायनिक बादल देने की अनुमति दी है। हमारी श्रद्धांजलि के पीछे, पुराने मध्य मैदान के दिमाग से जुड़कर केवल उस क्षण तक पहुंचा जा सकता है, क्योंकि उनके परिवर्तन के लिए, निर्माण जीव जैव रासायनिक प्रतिक्रियाओं (बी एन तारसोव, 1 9 74) के विकास में स्थिरता स्थापित करेगा। यह जीवन के स्थिर शिविर का मूल्यांकन करने और इसके संभावित नुकसान की भविष्यवाणी करने के लिए नई विधियों के विकास का भोजन बन गया है। महान लालच बायोथर्मोडायनामिक्स की शुरूआत और स्व-विनियमन प्रणालियों के साइबरनेटिक सिद्धांतों के जैविक अनुकूलन की प्रक्रियाओं के अनुवर्ती के बारे में है। यह स्पष्ट हो गया कि एक स्थिर राज्य की स्थिरता के बारे में भोजन को बढ़ाने के उद्देश्य से, तथाकथित कारकों की एक महत्वपूर्ण उपस्थिति, जो देखने के बिंदु पर, लिपिड ऑक्सीकरण की गैर-एंजाइमी प्रतिक्रियाओं को देखा जा सकता है। शेष घंटों में, डेडल जीवित कोशिकाओं के लिपिड चरणों में पेरोक्सीडेशन की आगे की प्रक्रियाओं और सक्रिय कट्टरपंथी उत्पादों के विकास का विस्तार करते हैं, जो झिल्ली के नियामक कार्यों को बाधित करते हैं। सक्रिय पेरोक्साइड रेडिकल्स की अभिव्यक्ति के रूप में प्रक्रिया के चक्र के बारे में जानकारी के स्रोत के रूप में सेवा करने के लिए, साथ ही पेरोक्साइड आधा जीवन (ए। टैपेल, 1965; आई। आई। इवानोव, 1965; ई। बी। बर्लाकोवा, 1967 और अन्य)। विकोरस रेडिकल्स की पहचान के लिए, बायोकेमोल्यूमिनेसिसेंस, जो उनके पुनर्संयोजन के दौरान जीवित कोशिकाओं के लिपिड के कारण होता है।
वाइनिकल्स की स्थिर अवस्था की स्थिरता के बारे में भौतिक और रासायनिक निष्कर्षों के आधार पर, एंटीऑक्सिडेंट सिस्टम के विघटन के रूप में बाहरी वातावरण के दिमाग के परिवर्तन के लिए विकास के अनुकूलन के बारे में बायोफिजिकल निष्कर्ष (बी। एन। तारसोव, हां ई. डोस्कोच, बी.एम. क्वेर्टला, 1968 - 1972)। इससे पाले और लवणता जैसी शक्ति का आकलन करना संभव हो गया, साथ ही कृषि और सूबेदार उत्पादकों के चयन के लिए भविष्य के पूर्वानुमानों में काम करना संभव हो गया।
1950 के दशक में, स्पेक्ट्रम के दृश्य और अवरक्त भागों (बी.एन. टारुसोव, ए.आई. ज़ुरावलेव, ए.आई. पोलिवोडा) में जैविक वस्तुओं की एक सुपरवीक लाइट - कम बायोकेमोल्यूमिनेशन थी। यह फोटोइलेक्ट्रॉनिक मल्टीप्लायरों (एल। ए। कुबेट्स्की, 1934) की मदद से सुपरवीक लाइट फ्लक्स को दर्ज करने के तरीकों के विकास के परिणामस्वरूप संभव हो गया। जीवित कोशिकाओं में होने वाली जैव रासायनिक प्रतिक्रियाओं का परिणाम होने के कारण, बायोकेमोलुमिनेसिस हमें एंजाइमों के बीच इलेक्ट्रॉनों के हस्तांतरण के लेंस में महत्वपूर्ण ऑक्सीकरण प्रक्रियाओं का न्याय करने की अनुमति देता है। वास्तव में, जैव रसायन विज्ञान का यह विकास महान सैद्धांतिक और व्यावहारिक महत्व का हो सकता है। तो, बी.एन. तारुसोव और यू.बी. कुद्रीशोव रोगविज्ञानी राज्यों के प्रतिशोध के तंत्र में गैर-आवश्यक फैटी एसिड के ऑक्सीकरण के उत्पादों की महान भूमिका को इंगित करते हैं जो कि कार्सिनोजेनेसिस और अन्य विकारों में आयनकारी इम्युनोसुप्रेशन के प्रभाव में विकसित होते हैं। सेल के सामान्य कार्य।
1950 के दशक में, परमाणु भौतिकी और बायोफिज़िक्स के अशांत विकास के संबंध में, रेडियोबायोलॉजी देखी गई, जो विप्रोमिनुवेन को आयनित करने की एक जैविक गतिविधि बनी रही। Obidna Prachny Radyoactive Izzotopiv, Bermonary Zbroї के सुसंगत, परमाणु ऊर्जा के परमाणु रिएक्टर व्यावहारिक वाइसरिस्टा के रूप, Zahista Svytaniva DIIA DIII ONZYUBOKO RADІYTSII, DIII ІONІZUKOїї Radіtskії, सैद्धांतिक बुनियादी बातों को लॉक करना। जिनके लिए हमारे लिए z'yasuvati का नमूना लेना आवश्यक था, क्लिटिन के याक घटक और भाषणों के लंका विनिमय सबसे अधिक समझदार हैं।
बायोफिज़िक्स और रेडियोबायोलॉजी के अध्ययन का उद्देश्य प्राथमिक रासायनिक प्रतिक्रियाओं की प्रकृति की खोज थी, जो उद्योग में ऊर्जा की वृद्धि के लिए जीवित सबस्ट्रेट्स को दोषी ठहराया जाता है। यहां इस घटना के तंत्र को समझना और भौतिक ऊर्जा को रसायन विज्ञान में बदलने की प्रक्रिया में बुद्धिमत्ता को जोड़ना, "भूरा" ऊर्जा के योग गुणांक को बदलना महत्वपूर्ण था। सोवियत संघ में एन.एन. सेमेनोव (1933) के स्कूल और इंग्लैंड में डी. हिंशेलवुड (1935) की विरासत से रोबोट सीधे तौर पर प्रेरित थे।
रेडियोबायोलॉजिकल अध्ययनों में एक महान स्थान ने विभिन्न जीवों के विकिरण प्रतिरोध का उच्च स्तर लिया है। यह स्थापित किया गया था कि बढ़ी हुई रेडियोरेसिस्टेंस (उदाहरण के लिए, ग्रिजली केलेट) लिपिड की उच्च एंटीऑक्सीडेंट गतिविधि द्वारा वातानुकूलित थी। क्लिटिन झिल्ली(एम। चांग टा इन।, 1964; एन। के। ओग्रीज़ोव टा इन।, 1969)। यह पता चला कि टोकोफेरोल, विटामिन के और थियोस इन प्रणालियों की एंटीऑक्सीडेंट शक्तियों के निर्माण में महत्वपूर्ण भूमिका निभाते हैं (आई। आई। इवानोव एट अल।, 1972)। शेष विश्व में, उत्परिवर्तजन के निम्नलिखित तंत्रों द्वारा स्वयं को बहुत सम्मान दिया जाता है। इस विधि का उपयोग इन विट्रो में न्यूक्लिक एसिड और प्रोटीन के व्यवहार के साथ-साथ वायरस और फेज (ए। गुस्ताफसन, 1945 - 1950) में आयनकारी प्रतिक्रियाओं को विकसित करने के लिए किया जाता है।
रासायनिक रक्षा की प्रभावशीलता में और सुधार के लिए संघर्ष, प्रभावी अवरोधकों की खोज और निषेध के सिद्धांत सीधे बायोफिज़िक्स के मुख्य कार्य हैं।
आगे बढ़ाया doslіdzhennya zbudzhennyh stanіv biopolimerіv, scho उनकी उच्च रासायनिक गतिविधि को दर्शाता है। सबसे सफल जागृति चरणों का विकास था, जिसे फोटोबायोलॉजिकल प्रक्रियाओं के पहले चरण के लिए दोषी ठहराया जाता है - श्रोणि के प्रकाश संश्लेषण।
इस प्रकार, रोसलिन पिगमेंट सिस्टम के अणुओं की प्राथमिक सक्रियता को समझने में महत्वपूर्ण योगदान दिया गया। अन्य सबस्ट्रेट्स पर सक्रिय पिगमेंट के खर्च के बिना वेक-अप स्टेशनों की ऊर्जा के हस्तांतरण (माइग्रेशन) के लिए एक महान मूल्य स्थापित किया गया है। इन घटनाओं के विकास में एक महान भूमिका ए.एम. टेरेनिना (1947 और बाद में) के सैद्धांतिक कार्य द्वारा निभाई गई थी। A. A. Krasnovsky (1949) ने क्लोरोफिल और योग एनालॉग्स के रिवर्स फोटोकेमिकल नवीनीकरण की प्रतिक्रिया की खोज की। नौ में एक भयंकर सामंजस्य है कि अगले एक घंटे में प्रकाश संश्लेषण को टुकड़ों में करना संभव होगा (div। भी विभाजित 5)।
बायोफिजिसिस्ट मियाज़ोवोगो गति की प्रकृति और तंत्रिका उत्तेजना और चालन के तंत्र की खोज पर काम करना जारी रखते हैं (डिव। धारा 11)। नबुली का वर्तमान महत्व जागृत अवस्था से आदर्श तक संक्रमण के तंत्र द्वारा भी पहुँचा जाता है। Zbudzheny को अब एक ऑटोकैटलिटिक प्रतिक्रिया और गैल्वनीकरण के परिणाम के रूप में माना जाता है - टोकोफेरोल (I. I. Ivanov, O. R. Kols6; 19 R. Kols6; 19 R. Kols6; 19 1970)।
बायोफिज़िक्स की सबसे महत्वपूर्ण जलती हुई समस्या जीवित पदार्थ की भौतिक और रासायनिक विशेषताओं की पहचान है। ऐसी शक्ति, जीवित बायोपॉलिमर के निर्माण के रूप में जो पोटेशियम को हिलाती है या विद्युत धारा का ध्रुवीकरण करती है, शरीर पर सुरक्षात्मक प्रभाव की उपस्थिति में बचाने की हिम्मत नहीं करती है। इसलिए, क्लिटिन बायोफिज़िक्स जीवित पदार्थों के अस्तित्व के लिए मानदंडों और विधियों का गहन विस्तार करना जारी रखता है।
मॉलिक्यूलर बायोलॉजी के यौवन में इसने मेरी नजर में जो सफलताएं हासिल की हैं, वे वाकई में चकाचौंध कर देने वाली हैं। अल्पावधि के लिए, जीन की प्रकृति और संगठन, कार्यान्वयन और कामकाज के मुख्य सिद्धांत स्थापित किए गए थे। उन पर, इन विट्रो में जीन के प्रजनन के रूप में zdіysnenno, और जीन का संश्लेषण पहले ही पूरा हो गया था। आनुवंशिक कोड को फिर से समझ लिया गया है और प्रोटीन जैवसंश्लेषण विशिष्टता की सबसे महत्वपूर्ण जैविक समस्या हल हो गई है। पता चला कि doslіdzheno कोशिकाओं में प्रोटीन की स्थापना के मुख्य पथ और तंत्र। समृद्ध परिवहन आरएनए की प्राथमिक संरचना, विशिष्ट एडेप्टर अणुओं को फिर से निर्धारित किया गया था, जिसके परिणामस्वरूप नए न्यूक्लिक टेम्पलेट्स को प्रोटीन के नए अमीनो एसिड अनुक्रम को संश्लेषित करने के लिए स्थानांतरित किया जाता है। अंत तक, समृद्ध प्रोटीन के अमीनो एसिड अनुक्रम को डिक्रिप्ट किया गया था और उनमें से एक विशाल संरचना स्थापित की गई थी। त्से ने एंजाइम अणुओं के कामकाज के सिद्धांत और विवरण की व्याख्या करना संभव बना दिया। एंजाइमों में से एक, राइबोन्यूक्लिएज के रासायनिक संश्लेषण पर काम किया गया है। विभिन्न उपचर्मीय कणों, समृद्ध विषाणुओं और फेजों के संगठन के मुख्य सिद्धांतों की स्थापना की गई, और भगशेफ में उनके जैवजनन के मुख्य तरीकों का अनुमान लगाया गया। Rozkryto जीवन के जीन और z'yasuvannya नियामक तंत्र की गतिविधि को विनियमित करने के तरीकों की समझ में जाते हैं। पहले से ही tsikh vіdkrittіv svіdchit की एक सरल रीटेलिंग, जो XX सदी का scho मित्र आधा है। जीव विज्ञान में एक महान प्रगति द्वारा चिह्नित किया गया था, जैविक रूप से सबसे महत्वपूर्ण मैक्रोमोलेक्यूल्स - न्यूक्लिक एसिड और प्रोटीन की संरचना और कार्य के मृत वृक्षारोपण से पहले एक प्रकार का गण्डमाला।
आण्विक जीवविज्ञान की उपलब्धियां आज पहले से ही अभ्यास में विजयी हैं और चिकित्सा, मजबूत राज्य और उद्योग की ड्यूस में फलदायी परिणाम देती हैं। निःसंदेह त्वचा दिवस के साथ विज्ञान का ज्ञान बढ़ता है। हालांकि, मुख्य विचार अभी भी विचार करने योग्य है कि, आणविक जीव विज्ञान में सफलता की आमद के साथ, जीवन के सबसे महत्वपूर्ण रहस्यों की खोज के मार्ग पर अकल्पनीय संभावनाओं की नींव पर जोर दिया गया था।
भविष्य में, शायद, पदार्थ के जीवन के जैविक रूप को प्राप्त करने के नए तरीके होंगे - जीव विज्ञान के आणविक स्तर से, परमाणु स्तर तक संक्रमण। हालांकि, एक ही समय में, शायद, भविष्य के उत्तराधिकारी जैसी कोई चीज नहीं है, ऐसा क्षण कि आणविक जीव विज्ञान के विकास को निकटतम 20 वर्षों में स्थानांतरित करना वास्तव में संभव था।
आणविक जीव विज्ञान / एम? मैंɛ इससे पहलेजेʊ मैंएर / gіlkoy जीव विज्ञान, scho डीएनए, आरएनए, प्रोटीन और उनके जैवसंश्लेषण के बीच की बातचीत के साथ-साथ इन अंतःक्रियाओं के विनियमन सहित, क्लिटिनी की विभिन्न प्रणालियों में जैव-अणुओं के बीच जैविक गतिविधि का आणविक आधार है। रिकॉर्ड प्रकृति 1961, एस्टबरी, आणविक जीव विज्ञान का वर्णन:
ऐसा तकनीकी नहीं है, जैसे कि pіdkhіd, pіdkhіd z pіdkhіd z pіdkhіd z pіdkhіd z pіdnіd z pіdіd z vіdnіvіn naukі isz provіdnoyu isdeєyu poshku कम बड़े पैमाने पर आणविक अभिव्यक्तियाँ klіhіdnogіd। विन टर्बोवानी टिम, ज़ोक्रेमा, सो फार्मजैविक अणु और [...] अधिक महत्वपूर्ण रूप से तुच्छ और संरचनात्मक रूप से - जिसका अर्थ यह नहीं है कि आकारिकी अधिक सटीक है। उस समारोह की उत्पत्ति को vodnochas doslіdzhuvati कर सकते हैं।
अन्य जैविक विज्ञानों तक रैंकिंग
आण्विक जीव विज्ञान के क्षेत्र में अनुयायी आणविक जीव विज्ञान को विकसित करने के लिए विशिष्ट तरीकों का उपयोग करते हैं, और अधिक से अधिक उन्हें आनुवंशिकी और जैव रसायन के तरीकों और विचारों के साथ जोड़ते हैं। मिज़ त्सिमी विषयों के बीच snuє नहीं सेवना। यह अगले आरेख में दिखाया गया है, जो खेतों के बीच पानी के एक संभावित दृश्य को दर्शाता है:
- जीव रसायन रासायनिक भाषणों का विकास और जीवों में होने वाली महत्वपूर्ण प्रक्रियाओं का जीवन। बायोकेमिस्ट्स के लिए बायोमोलेक्यूल्स की भूमिका, कार्य और संरचना पर ध्यान देना महत्वपूर्ण है। जैविक प्रक्रियाओं और जैविक रूप से सक्रिय अणुओं के संश्लेषण के लिए रसायन विज्ञान का अध्ययन जैव रसायन का अनुप्रयोग है।
- आनुवंशिकी जीवों में आनुवंशिक लक्षणों का संचार। यह अक्सर एक सामान्य घटक (उदाहरण के लिए, एक जीन) की उपस्थिति में पाया जा सकता है। तथाकथित "जंगली प्रकार" या सामान्य फेनोटाइप के संबंध में "उत्परिवर्ती" जीवों के प्रजनन में एक या अधिक कार्यात्मक घटक हो सकते हैं। जेनेटिक इंटरैक्शन (एपिस्टेसिया) अक्सर ऐसे "नॉकआउट" अनुक्रमों की सरल व्याख्याओं को भ्रमित करते हैं।
- आणविक जीव विज्ञान क्लिटिन की प्रतिकृति, प्रतिलेखन, अनुवाद और कार्य की प्रक्रियाओं के आणविक आधारों का विकास। आणविक जीव विज्ञान का केंद्रीय सिद्धांत यह है कि आनुवंशिक सामग्री को आरएनए में स्थानांतरित किया जाता है और फिर प्रोटीन में अनुवादित किया जाता है, चाहे क्षेत्र को समझने के लिए एक अच्छा प्रारंभिक बिंदु प्रदान करना कितना भी आसान क्यों न हो। आरएनए के लिए नई भूमिकाओं के आलोक में तस्वीर पर दोबारा गौर किया गया।
आणविक जीव विज्ञान के तरीके
आणविक क्लोनिंग
आणविक क्लोनिंग एक प्रोटीन के कार्य को बेहतर बनाने के लिए आणविक जीव विज्ञान में सबसे महत्वपूर्ण तरीकों में से एक है। इस तकनीक में, डीएनए जो रुचि के प्रोटीन को एनकोड करता है, उसे अतिरिक्त लैनजुग पोलीमरेज़ रिएक्शन (पीएलआर), और/या प्लास्मिड (एक्सप्रेशन वैक्टर) में प्रतिबंध एंजाइम के लिए क्लोन किया जाता है। वेक्टर 3 मान्यता प्राप्त चावल हो सकते हैं: एक प्रतिकृति सिल, और एक बहु क्लोनिंग साइट (MCS), एक चयनात्मक मार्कर, एक नियम के रूप में, एंटीबायोटिक दवाओं के लिए प्रतिरोधी। हमने प्रवर्तक क्षेत्रों के साथ कई क्लोनिंग के लिए एक साइट विकसित की है और दीक्षा की साइट के लिए प्रतिलेखन, जो क्लोन जीन की अभिव्यक्ति को नियंत्रित करता है। इस प्लास्मिड को बैक्टीरिया या पशु कोशिकाओं में डाला जा सकता है। जीवाणु कोशिकाओं में डीएनए की शुरूआत को नग्न डीएनए की मदद के लिए परिवर्तन के मार्ग से तोड़ा जा सकता है, अंतरकोशिकीय संपर्कों की मदद के लिए संयुग्मन, या वायरल वेक्टर की मदद के लिए पारगमन। भौतिक और रासायनिक गुणों की मदद से जीवित प्राणियों जैसे यूकेरियोटिक कोशिकाओं में डीएनए की शुरूआत को अभिकर्मक कहा जाता है। कई अलग-अलग ट्रांसफेक्शन विधियां उपलब्ध हैं, जैसे कैल्शियम फॉस्फेट ट्रांसफेक्शन, इलेक्ट्रोपोरेशन, माइक्रोइन्फेक्शन और लिपोसोमल ट्रांसफेक्शन। प्लास्मिड को जीनोम में एकीकृत किया जा सकता है, जिससे स्थिर ट्रांसफ़ेक्शन हो सकता है, या यह जीनोम से स्वतंत्र हो सकता है, जिसे ट्रांज़िशनल ट्रांसफ़ेक्शन प्रक्रिया कहा जाता है।
डीएनए, जिसे प्रोटीन सांकेतिक शब्दों में बदलना है, जो रुचि का है, वर्तमान में कोशिकाओं के बीच में है, और प्रोटीन अब व्यक्त किया जा सकता है। विभिन्न प्रणालियां, जैसे कि इंड्यूसिबल प्रमोटर और विशिष्ट सेल सिग्नलिंग कारक, उच्च स्तर पर प्रोटीन की रुचि को प्रेरित करने में मदद करते हैं। बड़ी मात्रा में प्रोटीन को बैक्टीरिया या यूकेरियोटिक कोशिकाओं के साथ जोड़ा जा सकता है। प्रोटीन विभिन्न स्थितियों में एंजाइमी गतिविधि से प्रभावित हो सकता है, प्रोटीन क्रिस्टलीकृत हो सकता है, इसलिए, ट्रेटिनस संरचना प्रभावित हो सकती है, या, दवा उद्योग में, प्रोटीन के खिलाफ नई दवाओं की गतिविधि प्रभावित हो सकती है।
पोलीमरेज़ लैनज़ग प्रतिक्रिया
मैक्रोमोलेक्यूल्स-ब्लॉट एंड रिसर्च
शर्तें pіvnіchny , ज़ाहिदनीі एक जैसाब्लॉटिंग इस तथ्य को ध्यान में रखता है कि आण्विक जीवविज्ञान गर्म है, क्योंकि यह शब्द पर था सदर्ननेट, एडविन सदर्न द्वारा BLOTTED डीएनए के संकरण के लिए वर्णित पद्धति के बाद पेट्रीसिया थॉमस, आरएनए रिटेलर - ब्लॉट, जो तब एक घरेलू नाम बन गया पिवनिचनी - सोख्तावास्तव में शब्द को हरा मत करो।
सदर्न ब्लॉट
योगी वाइनमेकर के सम्मान में नाम, जीवविज्ञानी एडविन पिवडेनी, फिर दक्षिणी - डीएनए नमूने में एक विशिष्ट डीएनए अनुक्रम की उपस्थिति पर आगे की जांच की विधि द्वारा धब्बा। अतिरिक्त जेल वैद्युतकणसंचलन के लिए पृथक्करण द्वारा प्रतिबंध एंजाइम (प्रतिबंध एंजाइम) को पचाने से पहले या बाद में डीएनए का विश्लेषण किया गया था, और फिर अतिरिक्त केशिका पाचन के लिए अतिरिक्त सोख्ता के लिए झिल्ली में स्थानांतरित कर दिया गया था। झिल्ली को तब एक लेबल डीएनए जांच के साथ अंतःक्षिप्त किया जाता है, जिसे डीएनए के अनुक्रम में जोड़ा जा सकता है, जो रुचि का है। डीएनए नमूनों से विशिष्ट डीएनए अनुक्रमों का पता लगाने के लिए पीएलआर जैसे अन्य तरीकों के विकास के कारण वैज्ञानिक प्रयोगशाला में दक्षिणी सोख्ता का व्यापक रूप से उपयोग नहीं किया जाता है। ये धब्बे अभी भी किसी प्रकार के शोध के लिए विजयी हैं, हालांकि, ट्रांसजेनिक चूहों में प्रतिलिपि संख्या ट्रांसजीन की कमी या भ्रूण स्टॉबर कोशिकाओं की नॉकआउट लाइनों के लिए जीन की इंजीनियरिंग में कमी।
पिवनिचनी ब्लोटिंग
उत्तरी धब्बा आरेख
सोख्ता के समान
आण्विक जीव विज्ञान से ज्ञात नैदानिक अध्ययन और उपचार के चिकित्सीय तरीके अक्सर जीन थेरेपी से दंग रह जाते हैं। चिकित्सा में आणविक जीव विज्ञान या आणविक क्लिटिन जीव विज्ञान दृष्टिकोण की स्थापना को अब आणविक चिकित्सा कहा जाता है। आण्विक जीव विज्ञान भी क्लिटिन के विभिन्न भागों की समझ, दीया और नियामक कार्यों को समझने में महत्वपूर्ण भूमिका निभाता है, क्योंकि इसका उपयोग नए चेहरों के प्रभावी संकेतों, रोग निदान और क्लिटिन के शरीर विज्ञान को समझने के लिए किया जा सकता है।
पढ़ने से दूर
- कोहेन, एसएन, चांग, एनकेडी, बॉयर, एच. एंड हेलिंग, आरबी जैविक रूप से कार्यात्मक जीवाणु प्लास्मिड का निर्माण नमूना पर .
1। परिचय।
विषय, आणविक जीव विज्ञान और आनुवंशिकी के तरीकों का कार्य। आणविक जीव विज्ञान और आनुवंशिक इंजीनियरिंग की स्थापना में "क्लासिक" आनुवंशिकी और सूक्ष्मजीव के आनुवंशिकी का महत्व। "शास्त्रीय" और आणविक आनुवंशिकी में एक जीन की अवधारणा, इसका विकास। आणविक आनुवंशिकी के विकास में आनुवंशिक इंजीनियरिंग की कार्यप्रणाली का परिचय। जैव प्रौद्योगिकी के लिए आनुवंशिक इंजीनियरिंग का अनुप्रयुक्त मूल्य।
2. क्षय की आणविक घात।
क्लिटिन के बारे में समझना, मैक्रोमोलेक्यूलर वेयरहाउस। आनुवंशिक सामग्री की प्रकृति। डीएनए के आनुवंशिक कार्य के प्रमाण का इतिहास।
2.1. विभिन्न प्रकार के न्यूक्लिक एसिड।न्यूक्लिक एसिड के जैविक कार्य। न्यूक्लिक एसिड की रासायनिक प्रकृति, अंतरिक्ष संरचना और भौतिक शक्ति। प्रोटो-यूकेरियोट्स की आनुवंशिक सामग्री की ख़ासियत। वाटसन-क्रिक फंडामेंटल्स की पूरक सट्टेबाजी। जेनेटिक कोड। आनुवंशिक कोड को समझने का इतिहास। कोड की मुख्य शक्तियां: ट्रिपल, कोमा के बिना कोड, वायरोजेनिटी। कोड डिक्शनरी की विशेषताएं, sim'ї kodonіv, smyslovі और "sily" kodonі। Kіltsі डीएनए अणु जो डीएनए सुपरस्पिरलाइज़ेशन के बारे में समझते हैं। उस प्रकार के डीएनए के टोपोइज़ोमर्स। डी टोपोइज़ोमेरेज़ के तंत्र। बैक्टीरिया का डीएनए गाइरेज़।
2.2. डीएनए प्रतिलेखन।प्रोकैरियोट्स के आरएनए पोलीमरेज़ में एक सबयूनिट और ट्रिविमिक संरचना होती है। सिग्मा कारकों की विविधता। प्रोकैरियोट्स, आईओगो संरचनात्मक तत्वों के जीन के प्रमोटर। ट्रांसक्रिप्शनल चक्र के चरण। दीक्षा, "समझा गया परिसर" का ज्ञान, प्रतिलेखन का विस्तार और समाप्ति। प्रतिलेखन क्षीणन। ट्रिप्टोफैन ऑपेरॉन अभिव्यक्ति का विनियमन। "रिबोर्मिक्स"। प्रतिलेखन समाप्ति तंत्र। प्रतिलेखन का नकारात्मक और सकारात्मक विनियमन। लैक्टोज ऑपेरॉन। लैम्ब्डा फेज के विकास में प्रतिलेखन का विनियमन। नियामक प्रोटीन (सीएपी प्रोटीन और लैम्ब्डा फेज रेप्रेसर) द्वारा डीएनए मान्यता के सिद्धांत। यूकेरियोट्स में प्रतिलेखन की विशेषताएं। यूकेरियोट्स में आरएनए प्रसंस्करण। प्रतिलेखों पर कब्जा, splicing और polyadeniation। स्प्लिसिंग तंत्र। छोटे परमाणु आरएनए और प्रोटीन कारकों की भूमिका। वैकल्पिक splicing, लागू करें।
2.3. प्रसारण, її चरण, राइबोसोम कार्य कोशिकाओं में राइबोसोम का स्थानीयकरण। प्रोकैरियोटिक और यूकेरियोटिक प्रकार के राइबोसोम; 70S और 80S राइबोसोम। राइबोसोम की आकृति विज्ञान। उप-भागों (उप-इकाइयों) में विभाजित। बढ़ाव चक्र में अमीनोसिल-टीआरएनए का कोडन-जमा बंधन। कोडन-एंटिकोडन इंटरैक्शन। राइबोसोम के लिए एमिनोएसिल-टीआरएनए के बंधन में बढ़ाव कारक ईएफ 1 (ईएफ-टीयू) की भूमिका। चिनिक बढ़ाव EF1B (EF-Ts), इसका कार्य, इसकी भागीदारी के साथ प्रतिक्रियाओं का क्रम। राइबोसोम से अमीनोसिल-टीआरएनए के कोडन-अपूर्ण बंधन के चरण में एंटीबायोटिक्स को जोड़ा जाना है। एमिनोग्लाइकोसाइड एंटीबायोटिक्स (स्ट्रेप्टोमाइसिन, नियोमाइसिन, केनामाइसिन, जेंटामाइसिन, आदि), संक्रमण का तंत्र। टेट्रासाइक्लिन अमीनोसिल-टीआरएनए के अवरोधक के रूप में राइबोसोम से जुड़ते हैं। प्रसारण दीक्षा। दीक्षा की प्रक्रिया के मुख्य चरण। प्रोकैरियोट्स में अनुवाद की शुरुआत: दीक्षा कारक, सर्जक कोडन, छोटे राइबोसोमल सबयूनिट का 3¢-टर्मिनल आरएनए और एमआरएनए में शाइन-डलगार्नो अनुक्रम। यूकेरियोट्स में अनुवाद की शुरुआत: दीक्षा कारक, सर्जक कोडन, 5¢-अनियंत्रित क्षेत्र और कैप-फॉलो "किंतसेवा" दीक्षा। यूकेरियोट्स में "आंतरिक" कैप-स्वतंत्र दीक्षा। ट्रांसपेप्टिडेशन। ट्रांसपेप्टिडेशन इनहिबिटर: क्लोरैम्फेनिकॉल, लिनकोमाइसिन, एमिसेटिन, स्ट्रेप्टोग्रामिन, एनिसोमाइसिन। स्थानान्तरण। बढ़ाव कारक EF2 (EF-G) और GTP की भूमिका। ट्रांसलोकेशन इनहिबिटर: फ्यूसिडिक एसिड, वायोमाइसिन, उनकी क्रिया का तंत्र। प्रसारण समाप्ति। टर्मिनुयुची कोडन। प्रोकैरियोट्स और यूकेरियोट्स को समाप्त करने वाले प्रोटीन कारक; समाप्ति के कारकों के दो वर्ग और dії के तंत्र। प्रोकैरियोट्स में अनुवाद का विनियमन।
2.4. डी एन ए की नकलऔर आनुवंशिक नियंत्रण। पॉलीमरेज़ जो प्रतिकृति में भाग लेते हैं, उनकी एंजाइमेटिक गतिविधियों का लक्षण वर्णन करते हैं। डीएनए पहचान की सटीकता। प्रतिकृति के घंटे के तहत डीएनए आधारों के जोड़े के बीच स्थैतिक अंतःक्रियाओं की भूमिका। पोलीमरेज़ I, II और III ई. कोलाई। सबयूनिट पोलीमरेज़ III। प्रतिकृति के घंटे के तहत विडेल्का प्रतिकृति, "प्रवीडना" और "चौड़ाई" धागे। टुकड़ा ओकाजाकी। प्रतिकृति कांटे पर गोरों का एक परिसर। ई. कोलाई प्रतिकृति दीक्षा का विनियमन। रोगाणुओं में प्रतिकृति की समाप्ति। प्लास्मिड प्रतिकृति के नियमन की विशेषताएं। उद्धृत किए जाने वाले रिंग के प्रकार के अनुसार द्विदिश प्रतिकृति और प्रतिकृति।
2.5. पुनर्संयोजन, ठेठ मॉडल। वैश्विक ची समजातीय पुनर्संयोजन। Dvonitkovі डीएनए विकसित करता है, scho innіtsіyuyut पुनर्संयोजन। डुप्लेक्स टूटने की प्रतिकृति के बाद की मरम्मत में पुनर्संयोजन की भूमिका। पुनर्संयोजन मॉडल में अवकाश संरचना। ई. कोलाई के वैश्विक पुनर्संयोजन का एंजाइमोलॉजी। आरईबीसीडी कॉम्प्लेक्स। रेका प्रोटीन। प्रतिकृति को बाधित करने वाले डीएनए क्षति के मामले में सुरक्षित डीएनए संश्लेषण में पुनर्संयोजन की भूमिका। यूकेरियोट्स में पुनर्संयोजन। यूकेरियोट्स में पुनर्संयोजन एंजाइम। साइट-विशिष्ट पुनर्संयोजन। वैश्विक और साइट-विशिष्ट पुनर्संयोजन के आणविक तंत्र का विवरण। पुनः संयोजक का वर्गीकरण। साइट-विशिष्ट पुनर्संयोजन के दौरान होने वाले गुणसूत्र परिवर्तन के प्रकार। बैक्टीरिया में साइट-विशिष्ट पुनर्संयोजन की नियामक भूमिका। साइट-विशिष्ट फेज पुनर्संयोजन की एक अतिरिक्त प्रणाली के लिए समृद्ध-कोशिका वाले यूकेरियोट्स के गुणसूत्रों का निर्माण।
2.6. डीएनए की मरम्मत।क्षतिपूर्ति के प्रकारों का वर्गीकरण। थाइम डिमर और मिथाइलेटेड ग्वानिन की सीधी मरम्मत। मूल बातें का विजन। ग्लाइकोसिलेज। बेमेल न्यूक्लियोटाइड की मरम्मत का तंत्र (बेमेल मरम्मत)। मरम्मत के लिए डीएनए स्ट्रैंड चुनें। एसओएस मरम्मत। प्रोकैरियोट्स और यूकेरियोट्स में एसओएस-मरम्मत में भाग लेने के लिए डीएनए पोलीमरेज़ की शक्ति। बैक्टीरिया में "अनुकूली उत्परिवर्तन" के बारे में कथन। जुड़वां किस्में की मरम्मत: डीएनए अणु के समरूप पोस्ट-रेप्लिकेटिव पुनर्संयोजन और बाद में गैर-होमोलॉगस छोर। प्रतिकृति, पुनर्संयोजन और पुनर्मूल्यांकन की प्रक्रियाओं की सहभागिता।
3. उत्परिवर्तन प्रक्रिया।
एक जीन - एक एंजाइम के सिद्धांत के निर्माण में जैव रासायनिक म्यूटेंट की भूमिका। उत्परिवर्तन का वर्गीकरण। बिंदु उत्परिवर्तन और गुणसूत्र परिवर्तन, उनकी स्थापना का तंत्र। सहज और प्रेरण उत्परिवर्तजन। उत्परिवर्तजनों का वर्गीकरण। उत्परिवर्तन का आणविक तंत्र। Vzaimozv'yazok उत्परिवर्तन और मरम्मत। म्यूटेंट की पहचान और चयन। दमन: आंतरिक, इंटरजेनिक और फेनोटाइपिक।
4. पॉज़ाक्रोमोसोमल आनुवंशिक तत्व।
प्लास्मिड, उनका भविष्य का वर्गीकरण। राज्य कारक एफ, योग जीवन और जीवन चक्र। क्रोमोसोमल ट्रांसफर को जुटाने में फैक्टर एफ की भूमिका। संयुग्म तंत्र। बैक्टीरियोफेज, उनकी संरचना और जीवन चक्र। विषाणुजनित और मृत बैक्टीरियोफेज। लाइसोजेनी और पारगमन। आईएस-एंबेसडरोवनोस्टे बैक्टेरियी, आईएस-एम्बुलाडोवनोस्टे याक घटक के एस-राजदूत एफ-फैक्टर बैक्टेरेयी, स्को वी ट्रांसपोसोनी ज़्दात्ने और कोनकोनि'एट बैक्टेरेई ताजियोटियन जीव ट्रांसपोसोनिव टीए रॉकी की संरचनाएं (एक्टोपिक रिकॉम्बिनेत्सिया) ये इवोलशिया जीनोम।
5. जीन की संरचना और कार्य पर शोध।
आनुवंशिक विश्लेषण के तत्व। सीआईएस-ट्रांस पूरकता परीक्षण। संयुग्मन, पारगमन और परिवर्तन की विविधताओं के साथ आनुवंशिक मानचित्रण। पोबुडोव आनुवंशिक कार्ड। सूक्ष्म रूप से आनुवंशिक रूप से मैप किया गया। जीन संरचना का भौतिक विश्लेषण। हेटेरोडुप्लेक्स विश्लेषण। प्रतिबंध विश्लेषण। अनुक्रमण विधि। पोलीमरेज़ लैंज़ुग प्रतिक्रिया। प्रकट जीन समारोह।
6. जीन अभिव्यक्ति का विनियमन। ऑपेरॉन और रेगुलेशन की अवधारणाएं। प्रतिलेखन की समान शुरुआत पर नियंत्रण कम है। प्रमोटर, ऑपरेटर और नियामक प्रोटीन। जीन अभिव्यक्ति का सकारात्मक और नकारात्मक नियंत्रण। प्रतिलेखन की समान शब्दावली पर नियंत्रण कम है। कैटोबोलिक-नियंत्रित ऑपेरॉन: लैक्टोज, गैलेक्टोज, अरेबिनोज और माल्टोज ऑपेरॉन के मॉडल। एटेन्यूएटर-नियंत्रित ऑपेरॉन: ट्रिप्टोफैन ऑपेरॉन का एक मॉडल। जीन अभिव्यक्ति का बहुविकल्पीय विनियमन। विनियमन की वैश्विक प्रणाली। तनाव के लिए नियामक प्रतिक्रिया। पोस्टट्रांसक्रिप्शनल नियंत्रण। संकेत पारगमन। आरएनए की भागीदारी के साथ विनियमन: छोटा आरएनए, सेंसर आरएनए।
7. जेनेटिक इंजीनियरिंग की मूल बातें। प्रतिबंध एंजाइम और संशोधन। उस जीन क्लोनिंग को देखकर। आणविक क्लोनिंग के लिए वेक्टर। पुनः संयोजक डीएनए के निर्माण के सिद्धांत और प्राप्तकर्ता कोशिकाओं में इसका परिचय। जेनेटिक इंजीनियरिंग के अनुप्रयुक्त पहलू।
ए)। मुख्य साहित्य:
1. वाटसन जे।, टूज जे।, रिकॉम्बिनेंट डीएनए: एक छोटा कोर्स। - एम .: लाइट, 1986।
2. जिन्न। - एम .: पवित्र। 1987.
3. आणविक जीव विज्ञान: न्यूक्लिक एसिड की संरचना और जैवसंश्लेषण। / ईडी। . - एम। विशा स्कूल। 1990.
4. , – आणविक जैव प्रौद्योगिकी. एम. 2002.
5. स्पिरिन राइबोसोम और प्रोटीन जैवसंश्लेषण। - एम।: आपका स्कूल, 1986.
बी)। डोडाटकोव साहित्य:
1. हेसिन जीनोम। - एम: विज्ञान। 1984.
2. रिबचिन जेनेटिक इंजीनियरिंग। - एसपीबी: एसपीबीजीटीयू। 1999.
3. जेनिव पेत्रुशेव। - एम: नौका, 2000।
4. आधुनिक सूक्ष्म जीव विज्ञान। प्रोकैरियोट्स (2 खंड में)। - एम .: लाइट, 2005।
5. एम। सिंगर, पी। बर्ग। जिनी और जीनोम। - एम .: लाइट, 1998।
6. शेल्कुनोवा इंजीनियरिंग। - नोवोसिबिर्स्क: जेड-वो सिब। विश्वविद्यालय।, 2004।
7. स्टेपानोव जीव विज्ञान। प्रोटीन की संरचना और कार्य। - एम: वी। श।, 1996।
साक्षात्कार
पिरोगोव सर्गी - 2012 में "हाथी और जिराफ" द्वारा आयोजित जीव विज्ञान में ओलंपियाड की तैयारी में भागीदार।
जीव विज्ञान के अंतर्राष्ट्रीय विश्वविद्यालय की उपलब्धि
ओलंपियाड "लोमोनोसिव" के विजेता
2012 में जीव विज्ञान में अखिल रूसी ओलंपियाड के क्षेत्रीय चरण के विजेता
एमडीयू आईएम से सीखें। एम.वी. लोमोनोसोव जीव विज्ञान के संकाय में: आणविक जीव विज्ञान विभाग, छठे वर्ष का छात्र। आणविक आनुवंशिकी संस्थान में जीवों के जैव रासायनिक आनुवंशिकी की प्रयोगशाला में अभ्यास करें।
- सर्जियस, अगर पाठकों के पास खाना है, तो क्या आप अपने ऊपर बदबू डाल सकते हैं?
तो, जाहिर है, आप एक बार में बिजली डाल सकते हैं। किसका क्षेत्र?
अनुरोध करने के लिए क्लिक करें।

- आइए इन स्कूलों को देखें, क्या आपके पास ऐसा स्कूल था जो सुपर कूल नहीं था?
मैंने मास्को के एक कमजोर स्कूल से सीखा, ऐसा औसत सांख्यिकीय स्कूल। सच में, हमारे पास MHK के एक चमत्कारी शिक्षक थे, zavdyaki yakіy हम स्कूल के नाममात्र "रहस्यमय ज्ञान" में बड़े पैमाने पर प्रकट हुए हैं।
- और जीव विज्ञान के बारे में क्या?
गर्मियों में हमारे द्वारा जीव विज्ञान का संचालन किया जाता था, एक बहरी और तेज औरत, उससे हर कोई डरता था। लेकिन प्यार ने को विषय नहीं दिया। खैर, मैं जीव विज्ञान के घुटन के बचपन से हूँ, पाँच के भाग्य। सब कुछ खुद पढ़ने के बाद, एनाटॉमी और जूलॉजी के लिए हांफना जरूरी है। बाद में, मेरे शक्तिशाली हितों के समानांतर स्कूल के विषयों की स्थापना की गई। Usy ने ओलंपियाड बदल दिया।
- मुझे अपनी रिपोर्ट के बारे में बताएं।
7 वीं कक्षा में, मैंने पहली बार नगरपालिका मंच का भाग्य लिया (मुख्य रूप से, एक बार फिर सभी विषयों से, उसी स्कूल के टुकड़े, जिसका पाठक प्रतिनिधित्व करने में सक्षम था)। मैं जीव विज्ञान में चैंपियन बन गया। तब स्कूल को कॉमेडी के समान स्तर पर रखा गया था, लेकिन उस cіkavogo तथ्य पर नहीं।

- आपने स्कूल में क्या मदद की?
मुझे याद है कि मुझे ब्लिसकुचा नवचन्न्या की परवाह नहीं है, अक्सर ओट्रिमुवव विद vykladach z biologії chetvіrki kshtalt पर prichіpkami के साथ "सिबुलिन रूट के एक छोटे से गुलाब के लिए, लेकिन rozfarbovani भूरा, सिरिम नहीं"। सभी ce bulo dosit टाइट। 8वीं कक्षा में, मैं फिर से ओलंपियाड में जीता, लेकिन जीव विज्ञान में उन्होंने मुझे सही नहीं किया। नेटोमिस्ट अन्य विषयों के विजेता और विजेता बने।
- और 9वीं कक्षा के बारे में क्या?
9वीं कक्षा में वे जिला स्तर तक नहीं पहुंचते। वहां मैं एक कमजोर, पास-पास की गेंद को स्कोर करने में विफल रहा, जो अभी भी क्षेत्रीय स्तर पर जा रही थी। मैं प्रेरक शक्ति को थोड़ा धक्का नहीं देता - मुझे पता है कि मैं नहीं जानता कि मैं कितना अमीर दिखता हूं और कितने अमीर लोग हैं, मुझे सब कुछ पता है (देश के पैमाने पर ऐसे कितने लोग हैं, मैं हूं प्रकट करने से डरते हैं)।

- मुझे बताएं कि आप कैसे तैयार हुए।
गहन स्वरोजगार, किताबों की दुकानों पर नबीगी और हजारों अन्य नौकरियों का बहुत कम प्रभाव पड़ता है। मैंने थ्योरी के लिए उच्चतम स्कोर में से एक स्कोर किया (मेरे लिए यह क्या समझना था), व्यावहारिक चरण में पास हुआ ... और योग में असफल रहा। तोडी मैं अभी भी व्यावहारिक चरण के औचित्य के बारे में नहीं जानने लगा।
- ओलंपिक ने आप पर क्या डाला?
मेरा जीवन मौलिक रूप से बदल गया है। मुझे कई अन्य ओलंपियाड के बारे में पता चला, विशेष रूप से ShPV से प्यार हो गया। एक वर्ष के लिए बैगातिओख पर अच्छे परिणाम दिखाने के बाद, विजयी रूप से "लोमोनोसिव्स्की" ज़ावड्याक्स जीता, बिना शराब के प्रवेश करने का अधिकार छीन लिया। उसी समय, मैंने कला के इतिहास में ओलंपियाड जीता, जिसके लिए मैं घबराहट से सांस लेता हूं और करता हूं। यह सच है, मैं व्यावहारिक दौरों के अनुकूल नहीं हूं। 11वीं कक्षा में, मैं अंतिम चरण में आगे बढ़ा, लेकिन फॉर्च्यून सुंदर नहीं था और मैं सैद्धांतिक चरणों के मैट्रिक्स को समझ नहीं पाया। लेकिन तब इसे प्रैक्टिकल के लिए अशांत नहीं होने दिया गया।
- क्या आपको अमीर ओलंपियाड के बारे में पता चला?
इसलिए, मैं इस बात का सम्मान करता हूं कि मैं अपने वन-लाइनर्स की हिस्सेदारी के साथ अधिक भाग्यशाली था, क्योंकि उन्होंने मेरे क्षितिज का काफी विस्तार किया। ओलंपियाड की दूसरी पुस्तक, प्रेरणा का अपराध अधिक सामंजस्यपूर्ण रूप से विषय को घुमा रहा था, यह ओलंपियाड से परिचित था। पहले से ही उस समय, मुझे याद आया कि ऊर्ध्वाधर के साथ घंटे का क्षैतिज संरेखण - शिविरों में vikladachs के साथ।

- आप वीएनजेड में कैसे शामिल हो रहे हैं? एक संकाय चुनना?
11वीं कक्षा के बाद मैंने एमडीयू के बायोलॉजिकल फैकल्टी में प्रवेश लिया। मेरे अधिकांश वर्तमान साथियों ने एफबीबी को कुरूपता के लिए घोटाला किया, लेकिन यहां पहली भूमिका उन लोगों द्वारा निभाई गई जो अखिल रूसी के विजेता नहीं बने। इसका मतलब है कि मुझे गणित के आंतरिक अनुभव को जोड़ने की आवश्यकता होगी, और इसमें, विशेष रूप से स्कूली बच्चों - मैं देख रहा हूं कि मुझे और अधिक प्यार हो गया है - मैं मजबूत नहीं हूं। स्कूल में, तैयारी कमजोर थी (हम महीने के अंत तक तैयार नहीं थे)। उसी समय, रुचियों की एक योजना होने के कारण, मुझे पता चला कि, मुझे यकीन है, आप किसी प्रकार का परिणाम प्राप्त कर सकते हैं, आवश्यकता की जगह की परवाह किए बिना। इन वर्षों में, यह स्पष्ट था कि बहुत सारे FBB स्नातक, याक एक बहुत ही महत्वपूर्ण जीव विज्ञान में उत्तीर्ण हुए, और दूसरी ओर - बहुत सारे अच्छे जैव सूचना विज्ञानियों ने शौकिया के रूप में शुरुआत की। हालाँकि उस समय मैंने सोचा था कि जैविक संकाय में FBBshny के बट में दल कमजोर नहीं होगा। Tsiomu में, मैं, bezperechno, दया करता था।
क्या तुम्हें पता था?
त्सिकावो
क्या तुम्हें पता था?
त्सिकावो
हाथी और जिराफ के शिविर में - जैव रसायन और आणविक जीव विज्ञान में परिवर्तन, एक ही समय में स्कूली बच्चे, एमडीयू के अंतिम रूप से, प्रयोगों की स्थापना करते हैं, और ओलंपियाड के लिए भी तैयार होते हैं।© साक्षात्कार ब्रावो रेशेतोव डेनिस। तस्वीरों ने कृपया पिरोगोव सर्गी को दबाया।




